Testando Seu Conhecimento

Aqui você encontrará uma série de questões que vão te auxiliar no aprendizado da matéria. Elas abordam cada tópico da aula dessa forma facilita sua fixação. E no fim você ainda pode verificar se suas respostas estão de acordo com o conteúdo. Para visualizar a resposta basta clicar na pergunta.

Membrana Plasmática
1 - Cite e explique 3 funções da bicamada lipídica:
- Envolver a célula, definir seus limites e manter as diferenças de concentração essenciais e movimento de solutos (por osmose ou por proteínas como carreadores e canais), devido à composição da membrana e sua propriedade semipermeável.
- Fazer síntese de ATP, pela migração de prótons de H+ através de uma proteína transmembranar na membrana mitocondrial interna (a ATP-sintase).
- Produz e transmite sinais elétricos e informações para o interior celular, permitindo respostas a variados estímulos (no caso de células musculares e nervosas), por meio de proteínas receptoras que captam moléculas sinais, na parte interna ou externa da membrana.
- Atua na comunicação entre células e compartilhamento de sinais que coordenam seu comportamento e expressão gênica, por meio de junções celulares.
- Atua na adesão celular, além de controlar orientação interna, movimento, crescimento e desenvolvimento celulares, por meio de filamentos do citoesqueleto ancorados entre si ou com a matriz.
2 - Diga a função do colesterol na membrana de células eucarióticas:
Ele modula as propriedades da membrana, se inserindo entre as caudas de hidrocarbonetos dos lipídeos e reduzindo sua mobilidade, tornando a membrana mais rígida e menos deformável nessa região, impedindo que as caudas se agrupem, e reduzindo sua permeabilidade a pequenas moléculas solúveis em água. Assim, torna a membrana mais fluida e menos suscetível à cristalização.
3 - Quais mecanismos a célula utiliza para evitar que sua membrana cristalize?
Cada célula passa do estado fluido para o cristalino ou gel (a “transição de fase”) a uma determinada temperatura; para se manter em meios com temperaturas variáveis, a membrana da célula é formada de uma variada mistura de fosfolipídeos, principalmente os com caudas mais curtas e com muitas insaturações (o que reduz a tendência das caudas de interagirem umas com as outras e evita que se agrupem), e a presença de colesterol garante mais rigidez e reduz o agrupamento das moléculas lipídicas. Isso permite que a membrana celular fique fluida em baixas temperaturas de transição de fase.
4 - Cite os dois tipos de proteínas de membrana e três diferenças entre elas:
Proteínas integrais:
- se inserem e se mantêm na região hidrofóbica da bicamada ou de uma monocamada;
- podem ser anfifílicas (possuem porções apolares e polares) e serem capazes de interagir com as porções polares e apolares das moléculas lipídicas e atravessar a membrana;
- podem ser solúveis e se ancorar a lipídios ou glicolipídeos por meio de ligações covalentes;
- podem adquirir forma de alfa-hélice ou barril-beta.
Proteínas periféricas
- não atravessam a membrana nem se estendem para a porção hidrofóbica da bicamada;
- ficam ancoradas em outras proteínas integrais;
- estão ligadas às proteínas por meio de ligações não covalentes, podendo se dissociar facilmente;
- podem se associar às proteínas do citoesqueleto e da membrana (dando forma à célula) ou da matriz extracelular (adesão a células ou substrato).
5 - Cite as semelhanças e diferenças na forma de difusão de moléculas lipídicas e de proteínas na bicamada:
Tanto fosfolipídeos como proteínas são capazes de se difundir lateralmente (trocando de lugar com outros lipídeos da mesma monocamada, no caso dos fosfolipídeos, e por dentro da membrana, no caso de proteínas). Enquanto os fosfolipídeos podem girar ao redor de seu próprio eixo, flexionar as caudas de hidrocarbonetos ou, mais raramente, migrar de uma monocamada para a outra (“flip flop”, que pode ser catalisado por proteínas como flipases, flopases e scramblases), as proteínas podem girar sobre um eixo perpendicular ao plano da membrana, mas não fazem flip flop.
6 – Defina glicosilação e descreva sua importância para a membrana celular executar algumas de suas funções essências:
A glicosilação garante o correto enovelamento da proteína, é uma reação química onde um carboidrato é adicionado a outra molécula e representa o ínicio da formação do glico-cálix. A glicosilação confere a membrana funções relacionadas a presença dos carboidratos ligados as proteínas , entre elas destacam-se a proteção , formação de uma malha de retenção de nutrientes e enzimas e possibilita o reconhecimento das células vizinhas.
7 – Esquematize e diferencie os 3 tipos de bombas de uma célula:
- Tipo P : bombeiam íons; são fosforiladas (mudam de forma) ATP = ADP + P
- Tipo F : bombeiam íons; produz ATP através do potencial de prótons (hidroelétrica na mitocôndria)
- Tipo ABC : transportam moléculas menores (aminoácidos, açucares, peptídeos etc); é conhecida como bomba de resistência as drogas (toxinas/proteínas ABC)
8 – Defina transporte ativo e transporte passivo estabelecendo diferenças entre eles:
- Transporte ativo: quando contra o gradiente eletroquímico, precisam de um fonte de energia, como por exemplo ATP, energia solar e gradiente eletroquímico (transporte acoplado)
- Transporte passivo: nos canais, sempre a favor do gradiente eletroquímico, também nos transportadores em alguns casos.
9 - Descreva de forma resumida o funcionamento e a importância da bomba de sódio e potássio para a célula:
A bomba de sódio e potássio bombeia sódio para fora e potássio para dentro da célula contra seus gradientes eletroquímicos. Para cada molécula de ATP hidrolisada dentro da célula, três íons de sódio são bombeados para fora e dois íons de potássio são bombeados para dentro, sendo assim é um processo ativo. Esta bomba é importante por estabelecer a diferença de carga elétrica entre os dois lados da membrana que é fundamental para as células musculares e nervosas e promove a facilitação da penetração de aminoácidos e açucares. Além disso, a manutenção de alta concentração de potássio dentro da célula é importante para síntese de proteína e respiração e o bombeamento de sódio para o meio extracelular permite a manutenção do equilíbrio osmótico.
10 - Descreva os componentes e como eles alteram as propriedades físicas da membrana plasmática.
A membrana plasmática é formada por uma bicamada lipídica e proteínas nela inseridos. Devido a natureza anfipática e a forma das moléculas lipídicas, as mesma formam bicamadas espontaneamente em solução aquosa. A membrana é um mosaico fluido que possui permeabilidade seletiva devido as características estruturais e químicas (cabeça hidrofílica e caudas hidrofóbicas) das moléculas lipídicas, tornando permeável a todas as moléculas hidrofóbicas e dificultando a passagem das demais substâncias sendo necessário algumas vezes de proteínas especializadas (bombas e canais) que facilitam o transporte das mesmas.
11 - Explique a característica de permeabilidade seletiva da membrana celular e que mecanismos ela utiliza para permitir a migração de moléculas através dela:
A membrana tem propriedade semipermeável, ou seja, as caudas de hidrocarbonetos dos fosfolipídeos que compõem a membrana atuam como uma barreira hidrofóbica; ela permite a passagem passiva de certas moléculas, como moléculas hidrofóbicas, e mais dificilmente de pequenas moléculas polares, mas impede a passagem de moléculas grandes ou carregadas. Estas moléculas podem atravessar a membrana com o auxílio de proteínas transmembranares; através de canais especializados, os quais permitem sua passagem passiva e a favor do gradiente de concentração, ou de transportadores, que gastam energia para atravessar as moléculas contra seus gradientes. Isso serve para manter um equilíbrio de concentração entre meios intra e extracelular, além de permitir a concentração intracelular adequada para o funcionamento da célula. Caso haja colesterol entre as caudas, essa permeabilidade diminui ainda mais.
12 - Explique a propriedade semipermeável da membrana das células e como elas a utilizam para se adaptar às diferenças de concentrações de soluto do meio extracelular.
As moléculas de água, embora polares, são pequenas o suficiente para atravessarem a membrana livremente; porém, moléculas como íons ou proteínas, que são carregadas ou grandes demais, não atravessam a membrana livremente, ficam retidas dentro ou fora da célula, e a concentração entre os meios intra e extracelular ficará desequilibrada. Para tentar manter a concentração equilibrada, as moléculas de água migram para o outro lado da membrana, por osmose. Se a concentração maior for a externa (solução hipertônica) elas migram para o meio externo e a célula tende a perder seu volume, correndo o risco de sofrer plasmólise; se for a interna (solução hipotônica), elas migram para o meio intracelular e a célula tende a ganhar volume, correndo o risco de sofrer lise.
Células animais (e algumas bactérias), por serem compostas apenas da membrana plasmática, são capazes de bombear os íons e outras moléculas através de bombas na membrana, para o exterior, para diminuir a concentração intracelular e evitar que se rompa; já células de plantas possuem ao seu redor uma parede celular rígida que permite que a célula tolere alterações nas concentrações, sem deformar a membrana, e que haja uma pressão interna capaz de expelir a água intracelular em excesso. No caso de protozoários, estes são capazes de formar e expelir vacúolos contrácteis com água.
Citoesqueleto
1 - Determine a função, localização e as principais funções do Microfilamento de Actina, dos Microtúbulos e Filamentos intermediários.
Microfilamento de actina: Formados por subunidades globulares proteicas de actina com um sitio de ligação de ATP ou ADP; Localizado geralmente próximo a membrana; Participa da contração muscular ; É responsável pela mobilidade da célula e participa da divisão celular; É uma estrutura altamente dinâmica (polimerização rápida dos microfilamentos de actina em determinada região).
Microtúbulos: Compostos por subunidades globulares e compactas , cada uma formada pelos monomeros alfa e beta-tubulina; Atravessam completamente a célula; Possui dois polos e uma origem em comum: o centrossomo; Formam os cílios e flagelos, participam do transporte de organelas e vesículas juntamente com proteínas motoras; Responsável pela segregação do DNA na divisão celular.
Filamentos intermediários: Formados por subunidades longas e fibrosas, existem diferentes tipos de filamentos, cada um constituído de moléculas específicas. Formam uma rede de filamentos no citoplasma; compõem o envelope nuclear dentre outras estruturas; Proporcionam força mecânica através de uma teia (cordas/filamentos); Responsável pelo ancoramento do núcleo e organelas.
2 - Relacione as colunas, preenchendo ou não todos os espaços com as consecutivas letras ou um x que representa a ausência de proteína motora correspondente:
A – Dineína
B – Cinecina
C – Miosina
( ) Microfilamentos de Actina
( ) Microtúbulos
( ) Filamentos Intermediários
3 - Cite 2 fatores que influenciam na instabilidade dinâmica do citoesqueleto.
A instabilidade dinâmica é um processo complexo e que depende de diversos fatores, entre eles podemos citar a concentração de subunidades livres no citoplasma ou na região alvo de formação específica no citoplasma. Também podemos citar a participação ou não de proteínas estabilizadoras, que mantém a forma e diminui a variação do citoesqueleto ligante.
4 - Diga quais são as subunidades que compõem cada um dos 3 tipos de filamentos de citoesqueleto.
Filamentos intermediários – Formados por subunidades longas e fibrosas, com porção central em hélice-alfa que forma uma estrutura paralela torcida com outra subunidade igual, formando um dímero. Ambos se associam com outro dímero, formando tetrâmeros; 8 tetrâmeros unidos formam uma secção com 32 hélices-alfa. Exemplos: queratina (queratinócitos); neurofilamentos (neurônios); laminina (lamina nuclear) e outros…
Filamentos de actina – Formados por subunidades globulares e compactas feitas de actina. A subunidade é uma estrutura globular protêica com um sitio de ligação de ATP ou ADP. Os monômeros se associam em oposição (cabeça e cauda) compondo filamentos com uma polaridade; um protofilamento se enrola sobre outro formam polímeros helicoidais de fita dupla organizados em feixes, redes ou géis.
Microtúbulos – Formados por subunidades globulares e compactas, feitas de tubulina. A subunidade é um heterodímero formado pelos monomeros alfa e beta-tubulina. Unem-se de forma longitudinal e lateral, formando longos cilindros ocos, rígidos e retilíneos, apresentando uma das extremidades ligada ao MTOC (centrossomo) que se expande para a célula.
5 - Identifique quais proteínas motoras e acessórias atuam no processo de contração muscular e explique este processo:
O Ca++ liberado no citosol se liga na troponina (acessória) presente nos filamentos finos provoca uma mudança em sua estrutura que acaba expondo o sitio de ligação da miosina nos filamentos de actina. Quando o ATP se liga à cabeça da miosina (proteína motora), ocorre uma mudança na conformação em que esta perde sua afinidade pela actina e, depois que o ATP é hidrolisado, a cabeça da miosina se eleva e liga-se a esse sítio ativo, tracionando o filamento espesso, feito de miosina, e fazendo-o deslizar sobre a actina, sem que seus tamanhos sejam diminuídos. A miosina é ancorados na linha Z através da proteína titina (proteína acessória), que age como uma mola e garante que o sarcômero volte à sua posição descontraída; quando ocorre a contração, a distancias entre as linhas diminui, aproximando as linhas Z, e o sarcômero encurta. A tropomiosina, presente nos filamentos finos, auxilia a manter sua estrutura contorcida.
6 - Cite 3 proteínas acessórias importantes para a polimerização de filamentos de actina e para microtúbulos:
Filamentos de Actina
- Complexo ARP – inicia a nucleação da actina
- Formina – acelera a polimerização da parte +
- Timosina – retarda o crescimento
Microtúbulos
- Gama-tubulina – inicia a nucleação das subunidades de tubulina
- Cinesina-13 – promove a rápida despolarização do microtúbulo
- MAP – estabiliza o microtúbulo e permite seu crescimento
7 - As proteínas acessórias Fimbrina (de filamentos intermediários) e TAU (de microtúbulos) auxiliam a formar a estrutura necessária de determinadas células para que estas exerçam suas funções. Diga qual a função dessas proteínas e em que células atuam:
Fimbrina – dá origem às microvilosidades, conectando as fibras entre si, em células intestinais.
TAU – mantêm a estrutura dos axônios, permitindo que ocorra o transporte de vesículas, em neurônios.
8 - Descreva a importância da presença dos filamentos intermediários de queratina para o tecido epitelial.
A queratina é sintetizada pelos queratinócitos, células diferenciadas do tecido epitelial. Devido à sua estrutura tridimensional, essa proteína possui propriedades particulares como impermeabilidade à água, alto nível de resistência e elasticidade. Essa proteína forma uma camada que envolve as células da epiderme (camada mais externa da pele), evitando perdas desnecessárias de água e, também, protegem o organismo contra agressões externas, tais como choques mecânicos, radiação solar, ventos e chuvas.
9 - Cite duas funções para cada proteína motora associada a microfilamentos de actina e ao microtúbulo.
Microfilamentos de actina
- Miosina I – organização intracelular e protusão de estruturas na superfície da membrana para formar microvilosidades.
- Miosina II – geração de força na contração muscular; liga-se às caudas de outras miosinas e sua cabeça hidrolisa ATP, usado como energia para que ela se prenda na actina e deslize sobre o filamento, tracionando-o.
- Miosina V – transporte de vesículas e organelas.
Microtúbulos
- Cinesinas – se movem sobre os microtúbulos (em direção à extremidade +); podem se conectar a uma organela delimitada por membrana ou outro microtúbulo; auxilia na formação do fuso mitótico ou meiótico e na separação de cromossomos durante a divisão celular.
- Dineínas citoplasmáticas – se desloca em direção à extremidade -; atuam no transporte de vesículas e posicionamento no Complexo de Golgi.
- Dineínas do axonema – atuam no rápido movimento de deslizamento dos microtúbulos para o batimento de cílios e flagelos (estes presos na superfície celular).
Junções Celulares
1 - Cite três funções das junções celulares:
- Manter as células coesas para formar estruturas multicelulares;
- Criar vias de comunicação que permitem o compartilhamento de sinais que coordenam o comportamento celular e regulam seu padrão de expressão gênica
- Controlar a orientação interna por meio de ligações a outras células e à matriz;
- Permitir que as células se movam e se desenvolvam, além de reparos celulares por meio da formação e quebra das ligações e da modelagem da matriz;
- Transmitir o estresse mecânico de uma célula para outra através de seus filamentos do citoesqueleto;
- Impermeabilização tecidual;
- Permitir a comunicação do citoplasma celular e, com isso, a troca de íons, pequenas moléculas e outros sinais.
2 - Quais os mecanismos que permitem que várias células sejam capazes de se unir de forma organizada e formarem os diferentes tecidos em um organismo multicelular?
A formação de um tecido se dá através da união das células entre si e com a matriz extracelular, devido às junções celulares que unem suas membranas e citoesqueleto, e através da transmissão do estresse entre essas células. Para isso, as células utilizam de duas construções diferentes, que variam em extensão ou componentes nos diferentes tecidos: a matriz extracelular (uma rede complexa de proteoglicanos) e os filamentos do citoesqueleto de cada célula, unidos de forma indireta pelas junções de ancoramento, que podem ser de adesão célula-matriz (como a adesão focal e os hemidesmossomos), ou de adesão célula-célula (como as junções aderentes e os desmossomos), que unem as células entre si e seu citoesqueleto.
3 - As junções de ancoramento são estruturas capazes de conectar as células entre si e transmitir uma força tensora entre todas elas, sendo muito abundantes em tecidos que sofrem grande estresse mecânico. Explique resumidamente quais são os tipos de junções de ancoramento, como as células interagem por meio dessa junção (cite as proteínas de adesão transmembranares) e com qual filamento do citoesqueleto elas se conectam:
- Junções aderentes – uma célula interage com outra célula por meio de proteínas da família das caderinas, que se ligam de forma homofílica entre seus domínios e indiretamente, por meio de proteínas adaptadoras (as proteínas de ancoramento), a filamentos de actina.
- Desmossomos – uma célula interage com outra célula por meio de proteínas da família das caderinas, que se ligam de forma homofílica entre seus domínios e indiretamente, por meio de proteínas adaptadoras (de ancoramento), a filamentos intermediários.
- Junções célula-matriz ou adesão focal – uma célula interage com a matriz extracelular ou lâmina basal por meio de proteínas da família das integrinas, que se ligam de forma heterofílica entre seus domínios e indiretamente, por meio de proteínas adaptadoras (de ancoramento), a filamentos de actina
- Hemidesmossomos – uma célula interage com a matriz extracelular ou lâmina basal por meio de proteínas da família das integrinas, que se ligam de forma heterofílica entre seus domínios e indiretamente, por meio de proteínas adaptadoras (de ancoramento), a filamentos intermediários.
4 - Quais são as principais proteínas participantes das Junções Ocludentes e qual sua importância?
As proteínas que participam destas junções são as claudinas, o principal componente, e as ocludinas, com função não determinada, que formam um “zíper” a partir da interação homofílica entre cada uma por toda a circunferência da célula. As Junções Ocludentes ou Tight Junctions são tipos de junções que ligam duas células e atuam como uma barreira de permeabilidade seletiva, separando os fluidos com composições químicas diferentes em cada lado. A interação entre as junções é extremamente fortes, sendo importante na manutenção da diferença essencial dos espaços corpóreos.
5 - DESCREVA Junções Comunicantes evidenciando suas subunidades, função e contextualizando sua importância na fisiologia humana.
As Junções Comunicantes (Junções Gap) ou nexos são formados por conexinas, proteínas transmembrânicas, que se unem formando o canal chamado conexon. São partículas cilíndricas que fazem com que as células entrem em contato umas com as outras, para que funcionem de modo coordenado e harmônico. Esses canais permitem o movimento de moléculas e íons, diretamente do citosol de uma célula para outra. A comunicação direta entres as células é muito importante para o funcionamento de diversas funções essenciais do nosso corpo, como a transmissão de sinais entre alguns tipos de neurônios através da passagem dos neurotransmissores por estes canais. A contração sincronizada do coração também depende da passagem ordenada de cálcio pelas junções comunicantes.
6 - DESCREVA o que é a Matriz Extracelular, suas funções e seus componentes fibrosos e fluídos.
Os tecidos não são constituídos apenas por células. Uma parte substancial do seu volume é de espaço extracelular que, por sua vez, é preenchido por uma rede de macromoléculas que constituem a MATRIZ EXTRACELULAR (MEC). Os componentes fibrosos se dividem em Fibrilares (colágenos fibrilares e proteínas de ligação) e Microfibrilares (colágeno IV, elastina e fibrilina). Já os componentes fluídos são representados pelos proteoglicanos (glicosaminoglicanos + proteínas). Essa Matriz extracelular e todos os seus constituintes executam várias funções, como preencher os espaços não ocupados pelas células, conferir aos tecidos resistência à compressão e ao estiramento, fornecer pontos fixos a diversos tipos de células para que elas possam se ancorar e possibilitar a rápida difusão de moléculas pelo tecido.
7 - DESCREVA o que é a lâmina basal, suas funções e todos seus principais componentes.
A Lâmina Basal é basicamente uma matriz extracelular em forma de placa fina e rígida constituída por colágeno tipo IV, perlecano e proteínas de adesão como entactina ou laminina. Está localizada em diferentes tecidos, como por exemplo abaixo de epitélios, nas células musculares, adipócitos, células de Schwann e glomérulo renal. Tem como função separar e prender o epitélio ao tecido conjuntivo adjacente, permitindo porém a passagem de diversas moléculas. A Lâmina Basal também determina a polarização da célula, influenciando sua diferenciação, proliferação, migração e morte.
8 - CITE qual a proteína mais abundante da matriz extracelular de origem animal e DESCREVA sua estrutura.
O Colágeno é a proteína mais abundante da matriz extracelular de origem animal e possui três cadeias α-hélice (tripla hélice) e cada cadeia é formada por repetições de sequências de aminoácidos: Gly-X-Y (glicina–prolina–hidroxiprolina).
Sistema de Endomembranas
1 - Descreva a diferença do RE das células musculares para células em geral e explique sua importância para contração muscular.
Diferentemente das células em geral, a célula muscular possui um tipo único de RE que apesar de não ter ribossomo em sua membrana não executa a função do RE Liso. O Retículo Sarcoplasmático é um RE específico das células musculares e executa funções essenciais para o processo de contração muscular. É nele que está armazenado os íons de cálcio, que induzem a Troponina a gerar uma mudança conformacional na Tropomiosina, que expõem os sítios de ligação da Actina para a Miosina se ligar e promover hidrólise gerando o encurtamento do sarcômero .
2 - Descreva a importância do processo de translocação co-traducional para a célula.
O processo de translocação das proteínas no Retículo Endoplasmático Rugoso é importante para a inserção da proteína na membrana e futura localização (membrana, vesículas, organelas como complexo de golgi e lisossomos, secreção). Esse processo também leva a glicosilação da proteína e controla seu correto enovelamento. A glicosilação é uma reação química onde um carboidrato é adicionado a uma molécula e representa o inicio do Glicocálix.
4 - O que são Polirribomos e porque eles são tão frequentes na célula?
A célula possui uma necessidade constante de proteínas para seu correto funcionamento, portanto ela precisa otimizar a produção das mesmas. Portanto, é possível e viável a tradução sequencial de um RNAm por vários ribossomos ao mesmo tempo, o que chamamos de polirribossomos. Este processo acelera a tradução e produção de proteínas.
5 - Descreva a Translocação co-traducional da proteína citando os elementos que atuam neste processo.
Após o inicio da tradução, se a proteína for direcionada à membrana do RER, dentre os primeiros aminoácidos estará presente uma sequência chamada de Sequência Sinal composta por uma sequencia de +-20 aminoácidos hidrofóbicos. A sequência é reconhecida pela SRP (Partícula Reconhecedora de Sinal), esta partícula possui uma cavidade de ligação à Sequência Sinal e quando é ligada a Sequência Sinal se dobra, conectando ao ribossomo seu domínio de pausa traducional, induzindo a pausa da tradução. A SRP será então reconhecida pelo receptor de SRP, na membrana do RER, encaminhando a proteína em formação para um translocador de proteína. A tradução contínua, após liberação da SRP do complexo, e a translocação se inicia, encaminhando a proteína em formação para o interior do RE rugoso. .
6 - Descreva quais os tipos de endocitose feitos pelas células:
Fagocitose – consiste na ingestão de partículas grandes ou de outras células (vírus ou bactérias) através de expansões citoplasmáticas do fagócito chamadas pseudópodos. Estes são capazes de detectar quimiotaticamente os micro-organismos (células responsáveis pela fagocitose possuem receptores de membrana que reconhecem substâncias específicas em bactérias) e depois os englobam, formando grandes vesículas endocíticas, os fagossomos, que irão se fundir posteriormente com os lisossomos. A fagocitose é dependente de reconhecimento por receptores de membrana.
Pinocitose – consiste na ingestão de fluidos e pequenas moléculas através de vesículas formadas na membrana plasmática, revestidas por clatrina ou caveolina, que formam fossas onde o líquido extracelular é aprisionado e internalizado, independendo de receptores de membrana.Os componentes endocitados são encaminhados para os endossomos. É um processo que ocorre continuamente nas células; a mesma quantidade de moléculas de membrana que são removidas com a pinocitose é devolvida por meio de vesículas secretórias, que se aderem à membrana, de forma que a área superficial da célula e seu volume permaneçam sempre constantes.
Endocitose – mediada por receptor consiste na ingestão macromoléculas concentradas em vesículas com clatrina que se ligam a receptores transmembrana complementares e acumulam-se em fossas, entrando na célula como complexos receptor-macromolécula dentro de vesículas. Os componentes endocitados são, então, encaminhados para os endossomos.
7 - Diga a função das proteínas de revestimento de vesículas, cite quais são elas, em quais regiões do tráfego de vesículas elas atuam (de onde se formam e para onde vão).
As proteínas de revestimento são capazes de se agregar em uma porção da membrana e deformá-la, interiorizando as substâncias próximas à membrana e dando forma à vesícula. As proteínas de revestimento se ligam a receptores de carga e, de acordo com o receptor, concentram substâncias específicas. As vesículas são utilizadas em diferentes etapas de seu transporte, dependendo de seu revestimento. Quando funcionam em diferentes locais ou vias elas normalmente incorporam subunidades proteicas que modificam suas propriedades.
São elas:
- Clatrina – em vesículas que brotam da membrana plasmática e levam material endocitado ou pinocitado desta para os compartimentos e para o complexo de Golgi;
- COPI – em vesículas que brotam do Complexo de Golgi para o Retículo Encoplasmático ou do Complexo de Golgi para a membrana plasmática;
- COPII – estritamente em vesículas que brotam do RE para a cisterna-cis do Complexo de Golgi.
8 - Diga o que são proteínas Rab e qual a função delas; exemplifique utilizando uma das proteínas Rab.
São proteínas endereçadoras, que sinalizam a origem da vesícula e as direcionam para seus locais específicos na célula. Para que as vesículas sejam fusionadas à membrana-alvo, esta precisa apresentar receptores para Rab complementares aos da vesícula a ela endereçada. Ex.: A proteína Rab1 está presente em vesículas que são formadas no Retículo Endoplasmático e no Complexo de Golgi, e são encaminhadas para as diferentes cisternas do Complexo de Golgi, pois estas possuem receptores de Rab1 em suas membranas; o receptor reconhece a Rab1 na vesícula, promovendo seu ancoramento.
9 - Cite os 2 tipos de fusão de membranas e diga como a fusão ocorre.
A fusão heterotípica ocorre entre a membrana de uma vesícula e a membrana de seu alvo, e a fusão homotípica ocorre entre as membranas de duas vesículas. A fusão entre as membranas ocorre através das proteínas de fusão, também chamadas de SNAREs. Existem dois tipos de SNAREs, as T-SNARE, presentes na membrana-alvo, e as V-SNARE, encontradas na membrana das vesículas. Essas proteínas possuem domínios helicoidais que, ao interagirem entre si, unem-se de forma estável e começam a se entrelaçar, formando o “complexo trans-SNARE”, que ancora as duas membranas. Esse entrelaçamento diminui a distância entre as membranas, expulsando moléculas de água existentes entre elas. Os lipídeos das membranas fluem de uma para outra, formando uma haste conectora; as camadas extracelulares das membranas se fusionam, e as membranas ficam “hemifusionadas”. Depois, as membranas internas se unem, ocorrendo a fusão completa.
10 - Diga quais as funções principais do revestimento feito nas vesículas de transporte pelas proteínas de revestimento (Ex. COPI, COPII e Clatrina):
- Concentrar proteínas específicas de membrana em uma região especializada da qual será formada a membrana da vesícula
- Selecionar moléculas-carga específicas para transporte através dos receptores de carga na membrana do compartimento doador.
- Modelar a vesícula em formação, se agregando em volta da membrana da vesícula e deformando-a, levando à envaginação da membrana e posterior liberação (brotamento) da vesícula da membrana original.
→ Uma vez feito esse processo, as proteínas de revestimento se soltam da vesícula já formada.
11 - As diversas hidrolases presentes no lisossomo funcionam apenas em meio de pH baixo (ácido), o qual a organela tende a mantê-las ativas e poder executar sua atividade de degradação. Por que essas hidrolases não funcionam no mesmo pH da célula (p. ex., o pH citosólico)?
Porque assim elas evitam digerir as proteínas da própria célula. Essas hidrolases são sintetizadas no citoplasma e passam pelo Retículo Endoplasmático e Complexo de Golgi; caso funcionassem no pH citosólico (em torno de 7,2), elas iriam degradar indevidamente as proteínas do citoplasma ou as presentes no interior dessas organelas antes mesmo de serem enviadas ao lisossomo. Outro motivo seria porque, caso haja um rompimento da membrana do lisossomo, essas hidrolases são incapazes de degradar as proteínas da célula porque permanecem inativas no pH citosólico.
Mitocôndria e Peroxissomos
1 - CITE 3 funções dos Peroxissomos.
Os peroxissomos são responsáveis por oxidar substratos orgânicos específicos (desintoxicação) , retirando átomos de hidrogênio e combinando-os com oxigênio molecular (O2). Essa reação resulta na produção de peróxido de hidrogênio (H2O2), uma substância oxidante muito prejudicial à célula. O H2O2 será rapidamente utilizado para oxidar outros substratos como fenóis, ácido fórmico, formaldeído e álcool. Essa reação, geralmente realizada pela enzima Catalese é útil para eliminar substancias tóxicas. O restante do H2O2 é imediatamente convertida em água e oxigênio também pela enzima catalase. Além disso, os mesmos são responsáveis por outro processo, a β-oxidação (transformação de ácidos graxos em acetil-CoA), que também pode acontecer nas mitocôndrias. Quando ocorre no Peroxissomo, a acetil-coenzima A (acetil-CoA), é enviado para o citoplasma celular. O acetil-CoA é utilizado em diversas reações de síntese e pode penetrar nas mitocôndrias para fornecer energia. Os peroxissomos possuem outras funções como, por exemplo, participar da síntese de ácidos biliares e de colesterol no fígado.
2 - DESCREVA o processo de formação dos Peroxissomos.
Os Peroxissomos se originam no Reticulo Endoplasmático através de vesículas precursoras. Estas vesículas apresentam proteínas específicas que catalisam a importação de proteínas e lipídeos do citosol. Todas as proteínas do Peroxissomo são importadas diretamente do citoplasma. A vesícula cresce devido a captação destes constituintes, que também provem sua diferenciação e aquisição de funções específicas.
3 - CITE a principal função da Mitocôndria e DESCREVA sua estrutura.
A principal função da Mitocôndria é de produção de ATP através da oxidação de acetil-CoA, originado da glicose, de lipídeos (β-oxidação ) ou proteínas. A Mitocôndria possui uma membrana externa, um espaço intermembrana, uma membrana interna e uma matriz mitocondrial. A membrana interna possui diversas invaginações que aumentam sua área, chamadas de cristas mitocondriais.
4 - Qual a importância da Mitocôndria apresentar dois diferentes espaços em sua estrutura?
O citosol possui uma baixa concentração de íons de hidrogênio (pH 7,2), e a produção de ATP na mitocôndria ocorre através da utilização da energia do gradiente eletroquímico destes íons pela ATP sintase. Portanto, seria impossível produzir ATP se não existisse um local na organela onde esse gradiente fosse presente e mantido. Este local é o espaço intermembrana, que possui uma alta concentração de íons de hidrogênio (pH 7,0). Essa diferença (gradiente eletroquímico) faz com que os íons passem para a matriz da mitocôndria, que é uma espaço com baixa presença de íons de hidrogênio (pH 7,5), através da ATP sintase e possibilita a formação de energia, utilizando a energia guardada no gradiente eletroquímico.
5 - DESCREVA como e porque a membrana interna da mitocôndria é diferente da membrana externa.
A membrana externa e a membrana interna da mitocôndria são bem diferentes , porém seu aspecto mais distinto pode ser considerado a permeabilidade. Um dos principais lipídeos que constituem a membrana interna é a cardiolipina, que possui 4 caudas de hidrocarbonetos responsáveis por diminuir bastante sua permeabilidade. Este aspecto é muito importante para o organela manter o gradiente eletroquímico imprescindível para a produção de ATP e as diferenças essências entre o espaço intermembrana e a matriz. Além disso, é na membrana interna da mitocôndria que se encontra a cadeia transportadora de elétrons e a ATP sintase, enzimas fundamentais para a respiração celular. Portanto, a membrana interna tem em sua composição 70% de proteínas, sendo a membrana com maior quantidade proteica da célula.
6 - Explique porque a respiração aeróbica é mais vantajosa à célula do que a anaeróbica.
Na respiração anaeróbica, é feita somente a glicólise, em que a glicose é transformada em duas moléculas de piruvato, com rendimento de apenas 2 ATPs e 2 NADH+ (depois o piruvato é transformado em lactato, na fermentação lática). Na respiração aeróbica, após conversão em piruvato, este segue para a mitocôndria, onde é transformado em Acetil-CoA, o qual entra no ciclo de Krebs, gerando um rendimento de 3 NADH +. Estes possuem elétrons altamente reativos que serão utilizados na cadeia transportadora de elétrons; cada elétron, após passar por seus complexos enzimáticos, irá gerar o bombeamento de 10 prótons (H+). No total, a respiração aeróbica gerará, com uma só molécula de glicose, de 30 a 32 moléculas de ATP, contra apenas 2 ATP somente da glicólise.
7 - Cite quais os dois principais substratos energéticos que podem ser utilizados pelas células. Qual possui um maior rendimento energético? Explique resumidamente como ocorre a obtenção de energia a partir de cada um.
A célula pode usar glicose ou, em certas ocasiões, os ácidos graxos, como substratos energéticos. A glicose pode ser transformada, através da glicólise, em duas moléculas de piruvato, gerando um saldo líquido de 2 ATPs e 2 NADH + H+; caso a glicose siga a via aeróbica, esse piruvato será encaminhado à mitocôndria, sendo transformado em Acetil-Coa. Já os ácidos graxos, na matriz mitocondrial ou no peroxissomo, sofrem β -oxidação, ou seja, uma clivagem a cada dois Carbonos em sua cauda de hidrocarbonetos, a qual pode variar em tamanho; o rendimento é de, no mínimo, 5 moléculas de Acetil-Coa e 5 NADH + H+. Na mitocôndria, o Acetil-Coa dos dois substratos pode ser utilizado na fase seguinte da respiração celular, o Ciclo de Krebs, em que sofrerá uma série de reações que terão como produtos um CO2 (expelido na expiração) e 3 NADH + H+, que permanecem no citoplasma. O NADH funciona como um aceptor temporário de elétrons; seu elétron altamente reativo será utilizado na cadeia transportadora de elétrons, resultando no bombeamento de vários prótons (H+) para o espaço intermembranar da mitocôndria, gerando um gradiente eletroquímico de H+ na membrana interna da mitocôndria, em que os prótons tenderão a voltar para a matriz e, para isso, irão passar pela bomba de prótons ATP sintase, a qual irá gerar ATP a partir de ADP e Pi a cada próton que retorna para a matriz.
No final, uma molécula de glicose, capaz de gerar dois Acetil-Coa, irá gerar de 30 a 32 ATPs, enquanto uma molécula de ácido graxo irá gerar três vezes mais, devido à formação de vários Acetil-Coa que irão entrar no ciclo de Krebs.
8 - Com o bombeamento de prótons para o espaço intermembranar da mitocôndria, durante o processo de respiração celular, é criado um gradiente eletroquímico. Explique seus dois componentes e como o gradiente eletroquímico pode auxiliar na produção de energia na célula.
O potencial de membrana (componente elétrico) torna a face voltada ao espaço intermembranar mais positiva do que a voltada à matriz, devido à carga positiva dos íons de Hidrogênio (pela polaridade, a tendência do H+ é voltar à matriz); o gradiente de concentração dos íons H+ (componente químico) torna-se maior no espaço intermembranar do que na matriz (logo, a tendência do H+ é também voltar à matriz, seguindo seu gradiente). Essas duas forças formam, então, um gradiente eletroquímico, e a tendência do próton é retornar à matriz; como a membrana interna da mitocôndria é impermeável a íons, o H+ não a atravessa livremente, mas é capaz de atravessar pela ATPsintase, localizada na mesma membrana. Os prótons de H+ passam por seu canal e, com isso, causam uma mudança de conformação em sua subunidade, que é capaz de “girar” e promover, na matriz mitocondrial, a síntese de ATP a partir do ADP e do Pi presentes na matriz.
9 - Onde as proteínas endereçadas para a mitocôndria são traduzidas e como elas são encaminhadas para ela?
Todas as proteínas são inicialmente traduzidas no citoplasma; no caso das proteínas direcionadas à mitocôndria elas estão totalmente traduzidas e livres no citoplasma, e possuem uma sequência sinal que a sinaliza para a mitocôndria. Essa sequência sinal é reconhecida por um receptor de sequência sinal de mitocôndria. Este receptor então encaminha a proteína a ser importada para um complexo proteico que catalisa a importação, localizado na membrana externa, chamado de Complexo TOM (Translocador da Membrana Externa). A proteína, então, atravessa a membrana externa e é capaz de ficar no espaço intermembranas ou pode ser reconhecida por outro complexo, o Complexo TIM (Translocador da Membrana Interna), na membrana mitocondrial interna, que logo capta a proteína e a atravessa por seu canal, inserindo-a na matriz mitocondrial. Na matriz, a proteína terá sua sequência sinal clivada por peptidases-sinal e adquirirá sua conformação, tornando-se uma proteína madura.
Núcleo Celular
1 - Diga as características de um poro nuclear e qual sua função.
Os poros nucleares são formados por diversas proteínas, as “nucleoporinas”, que formam uma simetria octagonal, e estão aderidos nas membranas do envelope nuclear. Ele é dividido em três porções: as fibrilas citoplasmáticas, projeções na face citosólica do envelope; cesta nuclear, as fibrilas que, na face nuclear, adquirem formato de cesta; e o cerne, porção transmembranar ou “corpo” que ancora o poro à membrana. Os poros conectam o citoplasma com o nucleoplasma e permitem um fluxo de moléculas nos dois sentidos (importação e exportação) ao mesmo tempo; essa passagem é feita rapidamente em moléculas com até 5kDa, ou em menor velocidade e com passagem regulada de 5kDa a 60 kDa; moléculas maiores do que 60kDa necessitam uma sequência sinal para sua passagem.
2 - A célula eucariótica possui, na membrana interna do núcleo, um revestimento de filamentos. Caracterize qual tipo de filamento do citoesqueleto é este, qual proteína a compõe e diga suas funções básicas.
Esse revestimento é a lâmina nuclear, formada por filamentos intermediários compostos de lamininas, responsáveis por garantir força mecânica e impedir que o envelope nuclear se desfaça; se relaciona com a cromatina e, durante o ciclo celular, as lamininas são fosforiladas, fazendo com que a lâmina nuclear se desfaça e causando a fragmentação do envelope nuclear. Isso permite a segregação dos cromossomos. As lamininas se desfosforilam ao final do ciclo celular e assim os vários fragmentos do núcleo se fusionam ao redor das cromátides condensadas, refazendo os núcleos das células-filhas.
3 - Imagine que, por alguma mutação, o núcleo de uma célula eucariótica não contenha nenhuma das estruturas subnucleares. Explique como ela seria prejudicada, dizendo qual a função dessas estruturas.
O nucléolo concentra os genes que codificam para os RNAr, responsáveis pelo processo de tradução de proteínas (junto com o RNAt), além da maquinaria para sintetizá-lo (RNA polimerase I, pré rRNA e pré-ribossomos); os Corpos de Cajal concentram a maquinaria responsável pelo processo de splicing, ou seja, o processamento de pré-RNA para RNAm (com a remoção dos íntrons), o qual possui a sequência para a tradução das proteínas. Logo, sem essa maquinaria devidamente concentrada, otimizando seus processos (síntese de RNAr e RNAm), a síntese de proteínas nessa célula seria prejudicada, ocorrendo mais lentamente.
4 - DESCREVA o processo de importação de proteínas no Núcleo.
Para uma proteína entrar no núcleo ela precisa conter em sua cadeia uma sequência sinal de localização nuclear (NLS). Esta sequência é facilmente reconhecida por um receptor de importação nuclear (impotina) que se liga a sua estrutura no citoplasma. Os receptores de importação carregados se ligam ao poro pelo lado citoplasmático e são movidos para dentro do núcleo. Em seu interior estão localizadas proteínas Ran ligadas à um GTP (Ran-GTP) que, ao se ligarem ao receptor importação carregado, liberam a proteína . O receptor de importação sem a presença da proteína volta para o citosol. Assim que o GTP na proteína Ran é hidrolisado no citosol, a mesma e o receptor de importação se desassociam para serem reaproveitados pela célula.
5 - Porque no núcleo encontramos regiões do DNA condensadas de maneiras distintas e como é possível essa condensação?
O material genético presente no núcleo é muito extenso, e nem todas a células possuem a mesma constituição e função, ou seja, nem todos os genes são expressos em todas as células. Portanto, surge a necessidade da cromatina possuir uma parte condensada que ocupa menos espaço e não transcreve RNA, a Heterocromatina, e possuir uma região que se encontra descondensada, podendo ser ou não ativa (ocorrendo ou não sua transcrição) que chamamos de Eucromatina. As principais proteínas responsáveis por essa diferenciação são as Histonas, que se unem ao DNA e se entrelaçam transformando uma estrutura chamadas de nucleossomos.
6 - DESCREVA o que é o Nucleossomo e como ele se apresenta no núcleo celular.
O Nucleossomo é composto por um pequeno segmento de DNA enrolado ao redor de um núcleo de histonas presente dentro do núcleo da célula. O Nucleossomo inclui aproximadamente 200 pares de nucleotídeos da fita de DNA, divididos em dois espirais, que se enrolam em torno de um disco proteico, constituído por quatro pares de proteínas chamadas histonas (H2A, H2B, H3 e H4). Dentre suas funções estão a condensação do DNA e a regulação de sua expressão.
7 - DESCREVA o processo de compactação da fita de DNA e as estruturas encontradas nos cromossomos:
A dupla-fita de DNA se entrelaça no complexo octamérico formados por proteínas histonas (4 pares de histonas), percorrendo duas voltas em cada uma delas (e assim formando uma unidade chamada nucleossomo), de forma contínua, formando então uma estrutura semelhante a um “colar de contas”, a cromatina, ou seja, o DNA mais as proteínas associadas, representadas principalmente pelas histonas. Dependendo da necessidade da célula, a cromatina pode se condensar em níveis variados. A eucromatina se encontra descondensada, com fibras de cerca de 11nm, podendo ou não ser ativa transcripcionalmente. Já a heterocromatina é altamente compactada, com fibras de cerca de 30nm e expressão inativada. Cromossomos condensados podem ter de 700nm (uma secção do cromossomo) até 1400nm (o cromossomo mitótico inteiro). O DNA compactado dos cromossomos possui regiões com sequências nucleotídicas específicas, além das informações genéticas armazenadas. A origem de replicação é uma região específica onde a maquinaria proteica relacionada à replicação da fita de DNA se liga para desfazer e abrir a dupla-hélice, permitindo o pareamento de nucleotídeos complementares aos da fita-molde. Os telômeros, localizados na extremidade das fitas, são diversas repetições de uma sequência de nucleotídeos determinada pela enzima telomerase, que os polimeriza, sendo não-codificante e servindo para manter a estabilidade estrutural do cromossomo, evitando a erosão de suas extremidades e uma consequente perda de informação genética. Já o centrômero é a região onde as duas cromátides-irmãs são unidas, formando um cromossomo. As duas cromátides são idênticas estruturalmente, surgindo da duplicação do cromossomo original, durante a intérfase, antes da divisão celular. Porém, durante este processo, as duas cromátides são separadas; o centrômero também possui estruturas proteicas que permitem que as fibras do fuso mitótico se conectem a cada cromátide. Ao serem tracionadas, as fibras as separam em direção a cada célula-filha.
DNA: Estrutura e Replicação
1 - DIFERENCIE e DESCREVA as diversas bases nitrogenadas presentes nos nucleotídeos.
As bases presentes nos nucleotídeos se dividem em dois grupos: pirimidinas e purinas. As pirimidinas são representadas por bases que contêm somente um anel de carbono com nitrogênio, classificadas em Uracila, Citosina e Timina. As purinas são representadas por bases que contêm dois anéis de carbono com nitrogênio e são divididas em Adenina e Guanina. No DNA estão presentes somente a Citosina que se une somente a Guanina e a Timina que se une somente com Adenina. No RNA estão presentes somente a Citosina, Guanina, Adenina e Uracila.
2 - DESCREVA com suas palavras o experimento realizado por Griffith em 1928 e continuado em 1948 por Avery–MacLeod–McCarty, e EXPLIQUE sua importância.
Para realizar o experimento Griffith utilizou duas bactérias parecidas, porém não iguais: CAP S (com cápsula; patogênica) e CAP R (sem capsula; não patogênica) e ratos de laboratório. Inicialmente ao introduzir a bactéria CAP S viva em um rato, após um período, o mesmo foi à óbito. E introduzindo a bactéria CAP R viva no rato, o mesmo permanece vivo. Ao aquecer a bactéria CAP S e promover a sua morte, Griffith percebeu que após inserido o rato permaneceu vivo. Entretanto ,ao misturar a solução de bactéria CAP s morta com a bactéria CAP R viva o resultado encontrado não foi esperado, ou seja, o rato após um período foi à óbito. Portanto, haveria algo na solução da bactéria CAP S morta que transformava a bactéria CAP R viva em patogênica, assim como a CAP S viva. Sendo assim, em 1948 outros pesquisadores Avery–MacLeod–McCarty separam a solução da bactéria CAP S morta por grupos representados por diferentes moléculas, como por exemplo um grupo de lipídeos, açucares e proteínas. E o resultado de óbito, ou seja, de transformação da bactéria CAP R viva em patogênica só ocorreu com a mistura da solução de ácidos nucleicos da bactéria CAP S morta. Assim, esses pesquisadores concluíram que os ácidos nucleicos (na forma de DNA) presentes no núcleo (material genético) são definitivamente os responsáveis por conter todas as informações genéticas das células. Portanto este experimento foi utilizado para comprovar a associação direta entre o material genético das células com suas características e hereditariedade.
3 - ESQUEMATIZE o Dogma Central da Biologia Molecular.
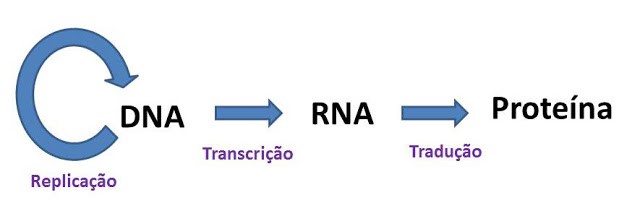
A Replicação, como o próprio nome diz, é o processo pelo qual o DNA é completamente duplicado gerando uma réplica, e este processo ocorre somente durante a divisão celular. A Transcrição representa a passagem das informações presentes em um gene (fragmento do DNA) para um RNA, para que o mesmo carregue as informações necessárias para a produção de uma proteína, ou seja, é o passo inicial para expressão genética (é uma ação constante para célula assim como a tradução) . A Tradução é o processo de decodificação dos nucleotídeos presentes no RNA em aminoácidos correspondentes, a cada três nucleotídeos é formado um códon que expressa um aminoácido específico. A união de vários aminoácidos através de ligações peptídicas, produz polipeptídeos e proteínas.
4 - Para que a fita-dupla de DNA mantenha sua conformação em hélice, dois diferentes tipos de interações são formados, entre diferentes nucleotídeos e diferentes bases nitrogenadas, uma intramolecular e outra intermolecular. Diga quais são essas ligações, onde são encontradas e qual a relevância de elas possuírem diferentes forças:
- Ligação de Hidrogênio (intermolecular): ligações relativamente fracas, encontrada entre bases nitrogenadas de nucleotídeos de duas fitas independentes de DNA (no “interior” de duas fitas). Variam em número de acordo com a estrutura da base; entre adenina e timina são formadas duas ligações e entre guanina e citosina, três (por isso, a ligação C-G é mais coesa do que a A-T).
- Ligação fosfodiéster (intramolecular): encontradas dentro de uma mesma fita de DNA, são ligações covalentes e, por isso, mais fortes; elas formam o arcabouço da fita de DNA, sendo encontradas entre o Carbono 3’ da desoxirribose de um nucleotídeo e o Carbono 5’ de outro. Na forma livre, a extremidade 3’ do nucleotídeo possui três fosfatos; dois deles são hidrolisados, encontrando-se na forma ionizada, e o fosfato restante compõe a ligação entre esses nucleotídeos.
- Pelo fato das ligações entre as duas fitas (pontes de hidrogênio) serem mais fracas que as ligações entre nucleotídeos da uma mesma fita (ligações covalentes fosfodiéster) é que a fita dupla de DNA pode ser despareada (desfeita ou separada) sem que haja ruptura da fita de DNA entre seus nucleotídeos, mantendo assim a informação de sequência de nucleotídeos intacta.
5 - Cite e explique quais os componentes necessários para que a replicação de DNA ocorra.
- Molde de fita do DNA, que contêm a sequência de nucleotídeos que servirá como base para a polimerização de uma fita de DNA complementar a ela.
- Trifosfato de desoxirribonucleotídeos livres (ATP; CTP; GTP; TTP) – para que sejam captados pela DNA polimerase e possam ser inseridos na cadeia em formação.
- Extremidade OH– livre em 3’ – para a incorporação de um trifosfato de desoxirribonuleotídeo, de acordo com a fita-molde; pode vir de um primer de RNA ou de um nucleotídeo já inserido.
6 - Cite as principais diferenças entre DNA e RNA:
DNA:
- Possui duas fitas de nucleotídeos, com pentoses e fosfatos dos nucleotídeos unidos, formando o “arcabouço” da cadeia, enquanto as bases nitrogenadas permanecem na parte interna, entre as duas fitas. As duas fitas enrolam-se uma ao redor da outra formando uma dupla-hélice.
- Nucleotídeos são Adenina e Guanina (purinas), Citosina e Timina (pirimidinas).
- O glicídio é uma desoxirribose sem hidroxila no Carbono 2’.
RNA:
- Possui uma única fita de nucleotídeos, os quais podem interagir entre si, conferindo à fita estruturas tridimensionais secundárias.
- É transcrita a partir do DNA; logo, seus nucleotídeos são complementares ao molde. São eles: Adenina e Guanina (purinas), Citosina e Uracila (pirimidinas).
- O glicídio é uma ribose com outra hidroxila no Carbono 2’.
7 - Cite três das principais enzimas ou complexos enzimáticos relacionados à replicação do DNA e descreva brevemente sobre seu funcionamento.
- DNA helicase: desfaz (despareia) a dupla-hélice nas fitas de DNA, conforme “caminha” pela cadeia, formando a forquilha de replicação.
- DNA primase: forma pequenos iniciadores (primers) de RNA na fita de DNA, na direção 5’-3’, os quais possuem uma extremidade 3′ livre para que a polimerização se inicie.
- DNA ligase: conecta os fragmentos de Okasaki após remoção de seus primers.
- Montador de cinta: complexo proteico que encaixa as cintas deslizantes (e, no caso dos fragmentos de Okasaki, também as remove para inserção de uma nova cinta a cada novo fragmento que será sintetizado). A cinta deslizante mantém a DNA polimerase firmemente associada ao DNA enquanto esta “caminha” pela fita.
- DNA topoisomerases: formam suportes giratórios no DNA e atuam de acordo com o tipo; a topoisomerase I liga-se em uma fita simples e cliva-a, permitindo que as duas fitas girem livremente, aliviando a tensão em pequenos segmentos da fita, e religando-a posteriormente; a topoisomerase II liga-se a duas duplas-hélices, clivando-as ao mesmo tempo e permitindo a passagem da cadeia através dessa abertura, religando-a depois.
- DNA polimerase: captam os trifosfatos de desoxirribonucleotídeos livres e catalisam a adição em sequência dos desoxiribonucleotídeos, fazendo a polimerização do DNA, efetuando a ligação fosfodiéster na extremidade OH– livre do nucleotídeo já inserido (ou do primer); remove os primers de RNA feitos para os fragmentos de Okasaki; através da atividade de correção exonucleotídica corrige nucleotídeos pareados incorretamente na extremidade 3’ da fita contínua, removendo-o em um sítio específico da enzima, regenerando a extremidade 3’ correta e refazendo a ligação com novos nucleotídeos.
8 - Cite e descreva brevemente quais as funções da DNA polimerase no processo de replicação do DNA.
A DNA polimerase é um complexo enzimático capaz de captar os trifosfatos de desoxirribonucleotídeos livres e catalisar a adição em sequência dos nucleotídeos, fazendo a polimerização do DNA e efetuando a ligação fosfodiéster na extremidade OH– livre do nucleotídeo já inserido (ou do primer). Ela remove os primers de RNA feitos no início dos fragmentos de Okasaki, para que estes depois sejam conectados por outra enzima, a DNA ligase. Através da atividade de correção exonucleotídica, ela corrige nucleotídeos pareados incorretamente na extremidade 3’ da fita contínua, removendo-o em um sítio específico da enzima, regenerando a extremidade 3’ correta e refazendo a ligação com novos nucleotídeos.
RNA a Proteína: Transcrição e Tradução
1 - DESCREVA os elementos fundamentais para que ocorra a transcrição e a função de cada um:
- Molde de DNA – chamada de fita codificante ou ativa, é o gene a ser transcrito e armazena o código genético.
- Enzima RNA polimerase – complexo enzimático responsável pelo pareamento de nucleotídeos livres ao molde de DNA e sua ligação para formação de uma fita de RNA. Quando associado a outras proteínas (fatores gerais de transcrição), é capaz de reconhecer a sequência de início de transcrição (TATA box), mover-se ao longo do DNA, despareando a dupla-fita e usando a fita codificante (o molde de DNA) para a transcrição do DNA em RNA, a fim de polimerizar uma cadeia de ribonucleotídeos complementar à fita codificante, assim como reconhecer a sequência de término da transcrição a fim de finalizá-la. Existem três diferentes tipos de RNA polimerase; I para sintetizar RNAr; II para sintetizar RNAm e III para sintetizar RNAt.
- Ribonucleotídeos trifosfatos livres – polimerizados em sequência formam a fita de RNA; possuem grupamento OH– no Carbono 2’ da pentose (diferente de deoxirribonucleotídeos) e são aderidos à fita de RNA em formação pela RNA polimerase, sendo as bases pareadas (Adenina, Uracila, Guanina e Citosina) complementares à fita codificante (C com G, G com C, A com T, e T com U).
2 - Cite e explique quais são as três etapas do processo de transcrição do RNA:
- Iniciação da transcrição – região promotora ou “promotor” é reconhecido por fatores gerais de transcrição, especificamente o TFIID, o primeiro a se conectar na dupla-fita; os demais fatores são recrutados depois, com uma ordem exata variando de acordo com o gene. Dessa forma, a RNA polimerase se conecta juntamente aos fatores e à dupla-fita, formando então o complexo de iniciação de transcrição, sendo capaz de atuar na transcrição. O TFIID junto à RNA polimerase separa a dupla hélice do DNA no ponto inicial de transcrição (após o promotor), expondo as bases da fita-molde do gene a ser transcrito, além de TFIID fosforilar a RNA polimerase, modificando sua estrutura e fazendo com que ela se desprenda dos demais fatores. A iniciação termina quando os primeiros dois nucleotídeos são ligados, passando então para a etapa de alongamento.
- Alongamento da transcrição – RNA polimerase, juntamente com o fator geral TFIID, se desprende dos demais fatores gerais de transcrição. O início da polimerização da molécula de RNA é marcado com a associação de pelos menos dois ribonucleotídeos. Dessa forma a ligação da RNA polimerase com a fita é estabelecida e permite a continuação do alongamento. Somente uma das fitas do DNA será usada como molde; o DNA vai sendo despareado à medida que a RNA polimerase se desloca sobre a fita enquanto sintetiza o RNA.
- Término da transcrição – essa etapa tem início quando a RNA polimerase reconhece uma sequência de terminação da transcrição. O RNA recém-formado sai pelo canal de saída de RNA da enzima RNA polimerase (ele vai se desligando do DNA que lhe serviu de molde conforme a enzima reconhece a sequência específica que indica o final do gene). Dessa forma, a transcrição é terminada e a RNA polimerase se desprende das fitas de DNA.
3 - Qual a principal diferença entre o processo de transcrição entre células procariontes para eucariontes? Diga de que forma essa diferença pode afetar a célula, levando em conta o fator evolutivo.
-
Células procariontes não apresentam um intermediário para a produção do mRNA a ser traduzido; o mRNA recém transcrito segue diretamente para a tradução (uma vez que a transcrição ocorre diretamente no citoplasma, devido à ausência de núcleo nessas células). Além disso, a maior parte do seu genoma é formada por sequências capazes de serem traduzidas para proteínas (regiões codificantes). Já em células eucariontes há a presença de um intermediário, o pré-mRNA, que é o RNA recém-transcrito e não modificado, ainda presente no núcleo. Ele então sofre as modificações pós-transcripcionais necessárias (splicing do RNA; capeamento e poliadenilação), tornando-se um mRNA maduro que seguirá ao citoplasma para ser traduzido. Apenas uma parte do genoma de células eucariontes pode ser traduzida para proteínas (parte que diminui conforme a complexidade do organismo aumenta; grande parte de nosso genoma é composto por genes não-codificantes).
4 - Existem três principais e abundantes tipos de RNAs envolvidos na tradução de proteínas. Diga quais são eles e como atuam, evidenciando o local de atuação.
- mRNA ou RNA mensageiro – fita simples de ribonucleotídeos; determina a sequência de aminoácidos em uma proteína após a tradução de seus códons em sequência de aminoácidos; marca o início e o fim da tradução com os códons de início (AUG) e parada posição (UGA, UAG, UAA), respectivamente; é formado de acordo com a necessidade da célula por sua proteína específica, sempre que necessário. No núcleo ainda pode ser encontrado como pré-mRNA e, após sofrer modificações específicas, segue ao citoplasma, onde será traduzido.
- rRNA ou RNA ribossomal – presente em maior quantidade, produzido no nucléolo (uma estrutura subnuclear do núcleo celular) e segue diretamente ao citoplasma, onde irá atuar; quando associados a proteínas são chamados ribossomos, que agem como sítios de tradução (são responsáveis pela síntese de proteínas a partir do código do mRNA maduro e dos aminoácidos carregados pelos tRNAs). O rRNA é capaz de efetuar a ligação peptídica entre o aminoácido trazido pelo tRNA e a cadeia de RNA em formação.
- tRNA ou RNA transportador ou de transferência – em menor quantidade; é produzido no núcleo, sendo transportado diretamente ao citoplasma, onde irá atuar; consiste em uma única cadeia de RNA que interage consigo mesma; dessa forma, possui duas regiões importantes: a extremidade 3`, o sítio de ligação ao aminoácido por onde é carregado o aminoácido específico ao códon do RNAm ou ao seu anticódon, complementar ao códon de RNA; e outra região, onde está localizada o anticódon, uma sequência complementar ao códon do mRNA e que pareia-se a ele para efetuar a ligação e reconhecimento do tRNA com o RNA e, consequentemente, a ligação do aminoácido na cadeia.
5 - Explique o que é um códon e um anticódon, e diga por que o código genético é dito “degenerado”.
Códons são sequências de três nucleotídeos específicos, presentes no mRNA, e que estão em sequência, a qual varia e, por isso, forma as diferentes informações genéticas a serem expressas e traduzidas. Sequências mais gerais marcam em que região do mRNA está o início da tradução da proteína (o códon de iniciação composto pelas bases AUG) ou o fim da tradução (códons de parada ou stop codon que são UGA, UAG, UAA, que não codificam a nenhum aminoácido). O anticódon é, também, uma sequência de nucleotídeos, porém está presente nos tRNAs e são complementares a seu códon (um códon do mRNA ACG, será pareado com um tRNA de anticódon UGC, por exemplo).
É devido ao fato de o códon ser específico e único para um aminoácido apenas (o qual é carregado pelo tRNA com anticódon complementar), mas um aminoácido ser codificado por um ou mais códons diferentes (assim como seus anticódons complementares), que o código genético é considerado “degenerado”. Através de um códon é possível ter certeza de seu aminoácido correspondente e da proteína que será formada; porém, um aminoácido pode ter sido codificado por diferentes códons, o que impossibilita o conhecimento exato do mRNA que a deu origem.
6 - Quais as principais modificações que o mRNA sofre antes que ele seja traduzido para proteína?
- Capeamento – o “cap” ou “quepe” consiste em um nucleotídeo guanina modificado (metilado) e “invertido” formado na extremidade 5’ do mRNA assim que ele emerge da RNA polimerase. O quepe é formado através da remoção de seu fosfato da extremidade 5’, seguido da adição de um GMP invertido (extremidades 5’ com 5’) e, por fim, a adição de um grupo metil a essa guanosina.
- Splicing – o processo de splicing ocorre após o capeamento do pré-RNAm. À medida que a RNA polimerase move-se no gene, o pré-RNAm sofre a remoção de seus íntrons (sequências não codificantes), feito por moléculas de RNA especializadas em um complexo chamado de spliciossomo, o qual é capaz de reconhecer as sequências entre dois éxons, onde o splicing deve ser realizado, a fim de juntá-los. Os dois éxons são unidos com a remoção do íntron entre eles, como um “laço”.
- Poliadenilação – após o término da transcrição e a formação de um pré-mRNA ocorre a formação da cauda poli-A na extremidade 3’ do mRNA pela enzima poli-A-polimerase, que adiciona até 200 nucleotídeos de Adenina à extremidade 3’ do mRNA. A extremidade final do mRNA possui a sequência de término da transcrição e adição de cauda poli-A, a qual é reconhecida por duas proteínas específicas que se ligam a ela e adicionam sequências específicas para “criar” a extremidade 3’, garantindo um grupo OH– em 3’. O tamanho final dessa cauda é ditado por proteínas que se ligam a ela conforme é sintetizada. Após o término da poliadenilação, o mRNA pode ser dito como um “mRNA maduro”.
7 - DESCREVA os elementos fundamentais para que ocorra a tradução e a função de cada um.
Os elementos fundamentais para que ocorra a tradução são: aminoácidos ligados a RNAt (aminoacil-tRNA), ribossomo e RNA-m. O RNAt através de seus anticódons pareia com os códons presentes no RNAm e transporta o aminoácido correspondente para a síntese, dentro do ribossomo. O RNAt representa o papel de um interprete (adaptador) entre a “linguagem” do RNA-m e a das proteínas. É o RNAm quem carrega todas as informações presentes no gene que foram transcritas no núcleo e exportadas para o citosol. O Ribossomo é a maquinaria composta por RNAr e proteínas associadas responsável pela união dos diferentes aminoácidos, carregados e selecionados pelos RNAt ,através de ligações peptídicas, formando assim as proteínas.
8 - DESCREVA resumidamente as etapas da tradução.
A tradução possui três etapas: iniciação, alongamento e termino. A tradução se inicia com a ligação da subunidade pequena do ribossomo ao início do RNAm. A subunidade pequena do ribossomo então varre o RNAm até encontrar o primeiro códon de iniciação (AUG). O reconhecimento do códon iniciador é feito pelo RNAt iniciador que possui o anticódon UAC e que transporta o aminoácido metionina. Quando ocorre a ligação do RNAt iniciador com o códon de iniciação esse complexo recruta a ligação da subunidade grande do ribossomo. Agora o ribossomo se encontra completamente montado e fixado, apresentando quatro sítios: A, P, E e o sítio de interação com RNAm. O complexo RNAt + aminoácido possui o nome de Aminoacil-tRNA. Ele entra através do sitio A (sitío aminoacil-tRNA) do ribossomo e pareia com as bases presentes no RNAm através do pareamento do anti-códon do RNAt com o códon no RNAm, carregando consigo um aminoácido correspondente. Com a continuidade da tradução este Aminoacil-tRNA é movido para o sítio P. Um segundo aminoacil-tRNA entra pelo sítio A, através do pareamento códon/anti-códon. Neste momento o ribossomo catalisa a união dos aminoácidos (um presente no sítio P e outro presente no sítio A) através da formação de uma ligações peptídicas, produzindo um Peptidil-tRNA que se encontra no sítio A. Esse complexo (peptidil-tRNA) é então movido para o sitio P do ribossomo (sítio peptidil-tRNA) e o então vazio sítio A recebe uma nova molécula de aminoacil-tRNA. Ao entrar diferentes Aminoacil-tRNA no sitio A e prosseguir a síntese proteica , RNAt vazios saem através do sítio E (de EXIT, saída) e aminoácidos recém ligados saem do ribossomo por um orifício. Essa etapa onde ocorre a síntese proteica é chamada de alongamento. Assim que o RNAm apresentar o códon de terminação (Stop códon, UGA, UAG, UAA) nenhum RNAt é correspondente a essa sequência, portanto, uma proteína chamada fator de liberação pareia com este sinal e assim que entra em contato com o sítio P no ribossomo provoca a dissociação da maquinaria e a consequente parada da Tradução, liberando a proteína traduzida. Etapa denominada, portanto, de termino da tradução.
9 - O que é e qual a importância da Aminoacil-tRNA-Sintetase?
A Aminoacil-tRNA-Sintetase é uma enzima responsável pela união do RNAt e do aminoácido correspondente ao seu anti-códon para formar um Aminoacil-tRNA específico. Portanto, existem pelo menos uma para cada aminoácido. Como o código genético é degenerado (a maior parte dos aminoácidos são codificados por mais de um códon diferente), em geral existe uma Aminoacil-tRNA sintetase para cada códon (ou entorno de 61 enzimas). Essa enzima reconhece o aminoácido e o anti-códon, promovendo a união e a montagem de diversos Aminoacil-tRNA essenciais para o processo de tradução, produção de proteínas e consequente execução de quase todas as funções celulares.
10 - ESQUEMATIZE e DESCREVA todas as características do Código Genético.
- Universalidade do Código Genético: códons expressam o mesmo aminoácido na maioria dos seres vivos.
- Não ambiguidade: cada códon codifica apenas UM aminoácido.
- Degenerado: códons diferentes podem codificar o mesmo aminoácido.
- Códon de terminação: os códons UAA, UAG e UGA terminam a síntese da proteína e não codificam nenhum aminoácido .
- Códon de iniciação: códon AUG que apresenta dupla função: codifica a Metionina e inicia a leitura do código.
11 - Como é possível uma sequência linear de nucleotídeos (RNA) com apenas 4 diferentes letras ser traduzida para 20 diferentes tipos de aminoácidos?
A resposta é bastante simples e pode ser encontrada no código genético, que corresponde ao dicionário que a célula utiliza para traduzir a linguagem genética em linguagem proteica. Apenas quatro letras não poderiam codificar todos os aminoácidos existentes, sendo assim, a célula utiliza uma sequência de três letras (três nucleotídeos), que representa um códon, para codificar um aminoácido específico. Deste modo, existem até mais possibilidades de codificação que aminoácidos existentes, 64 possibilidades e 20 aminoácidos existentes.
12 - Existe diferença entre o início de tradução de proteínas entre eucariotos e procariotos?
Sim. Em procarioto o códon de início de tradução é representado pelo primeiro AUG após sequências específicas do RNAm que se ligam ao ribossomo, chamadas de Sequencia de Entrada de Ribossomo (RES). E em eucariotos o códon de início de tradução é o primeiro AUG após o CAP presente no início do RNAm.
Ciclo Celular: Divisão Celular, Mitose e Meiose
1 - CITE qual complexo induz a entrada da célula na fase M e DESCREVA sua ação.
O complexo M-Cdk induz a entrada da célula na fase M do ciclo celular. Portanto, a ação do mesmo está diretamente relacionada ao efetivo processo mitótico, atuando na ativação de diversas proteínas e estimulando muitos eventos necessários, como a fosforilação das lamininas que são filamentos intermediários e compõem a lâmina nuclear, causando sua fragmentação e o desaparecimento do envoltório nuclear; a fosforilação da condensina que promoverá a condensação dos cromossomos e das coesinas que levará a separação das cromátides irmãs; e a fosforilação de proteínas associadas aos microtúbulos, importantes para a formação do fuso mitótico.
2 - DESCREVA a divisão do Ciclo Celular e as subdivisões da fase M.
O Ciclo Celular foi divido em 4 fases sequenciais: fase G1, fase S, fase G2 e M. Cada fase possui uma característica peculiar: A fase G1 (ou fase de intervalo 1; do inglês GAP) inicia-se logo após o fim da divisão celular anterior e nesta fase as células-filhas crescem, aumentam de volume, sintetizam membrana e organelas novas para adquirir volume semelhante ao da célula-mãe, a partir disso podemos concluir que a maioria das células se encontra nessa fase. Na fase S o DNA é duplicado, e assim, denominada a fase de síntese. A fase G2 pode ser considerada uma segunda fase de crescimento em que ocorre o preparo para a divisão celular e a analise se o DNA foi corretamente replicado e se, portanto, não possui danos em sua estrutura. Na fase M o material genético duplicado é dividido para duas células-filhas e é considerada uma fase de maior complexidade, deste modo, a mesma é subdivida em Prófase, Prometáfase, Metáfase, Anáfase, Telófase e Citocinese, possuindo cada subfase características especificas.
3 - Quais as diferenças entre uma célula haploide e uma célula diploide.
Células haploides possuem apenas uma cópia de cada gene, podendo ser organismos completos ou serem gametas de organismos diploides, que são as células responsáveis pela reprodução sexuada e que após a fecundação manterá o seu genoma em um tamanho original (diploide). Uma célula diploide possui duas cópias de cada gene, assim como todas as células do nosso corpo (com exceção dos gametas). Para a maioria das células do nosso corpo possuir um estado haploide é preciso inicialmente portar um estado diploide para que a divisão desta célula primordial desenvolva duas células com a metade de seu genoma. Esta divisão é denominada meiose, que representa a divisão celular especializada para a geração dos gametas. A divisão celular que mantém constante o tamanho do genoma, ou seja, produz células diploides é denominada mitose. Entretanto, é preciso lembrar que existem seres haploides, que não realizam a meiose e portanto não produzem gametas. O essencial é saber que a mitose mantém a ploidia da célula e a meiose reduz a ploidia a metade, podendo formar gametas ou não (plantas fazem meiose para produzir esporos que não são gametas).
4 - O que é e quando ocorre a clivagem.
A clivagem é um tipo diferente de ciclo celular, em que ocorrem diversas divisões sem crescimento e sem restabelecimento do corpo celular. Este processo ocorre durante o desenvolvimento embrionário e demonstra como um organismo multicelular surge através de uma série de divisões mitóticas, a partir de um grande volume de citoplasma do embrião que é dividido em numerosas e pequenas células nucleadas, chamadas blastômeros. O zigoto é dividido inicialmente ao meio, depois em quartos, oitavos, e assim por diante. Essa divisão sem crescimento do volume citoplasmático ocorre pela ausência das fases G1 e G2 da mitose, entretanto a divisão nuclear ocorre em níveis extremamente altos.
5 - Como é realizado o controle e a manutenção do Ciclo Celular?
O controle e a manutenção do ciclo celular são realizados através dos pontos de verificação e é a ligação das CICLINAS específicas com as CDK que indicam a passagem para próxima etapa, ou seja, a continuidade correta do ciclo. A Checagem de iniciação (G1/S) analisa se há condições ambientais para a célula se proliferar (nutrientes, fatores de crescimento), a Checagem da entrada em Mitose (G2/M) verifica se não há danos no DNA e a Checagem para a anáfase e citocinese se há problemas na formação do fuso mitótico e consequente problemas na segregação do DNA. Esse mecanismo funciona como um relógio que liga cada evento em um tempo específico e faz com que os mesmos sejam sempre ordenados. Como são acontecimentos cíclicos, essa verificação garante que cada evento ocorra somente uma vez e que as etapas sejam realmente finalizadas antes do inicio da próxima. Apesar de ser claramente um processo complexo e muitas vezes eficiente, alguns erros podem acontecer. É um processo facilmente adaptável à tipos celulares específicos ou condições ambientais adversas.
6 - Qual a função da Condensina?
Como o próprio nome induz, a Condensina contribui para a espiralização e condensação do DNA na fase M.
7 - Qual complexo garante que o DNA seja replicado apenas uma vez?
O Complexo pré-replicativo se forma no início da Fase G1 e garante que o DNA seja replicado apenas uma vez, ligando-se às origens de replicação.
.
8 - Diga qual a diferença entre os processos de mitose e meiose, evidenciando as funções desses processos.
Na mitose o tamanho do genoma sempre permanece constante na divisão (uma célula diploide terá células-filhas diploides); esse tipo de divisão é essencial para a proliferação, tanto para seres unicelulares (procariontes, como bactérias, por exemplo) como para os multicelulares, os quais são formados a partir de uma única célula em constante divisão, formando os diferentes tecidos conforme suas células vão se diferenciando e se especializando. Na meiose, o genoma é dividido entre as células-filhas com metade de sua ploidia (uma célula diploide terá duas células haploides); geralmente são células localizadas em órgãos especializados, os órgãos reprodutores, e são responsáveis pelas formação de gametas haploides (espermatozoides e ovócitos).
9 - A seguir estão, em sequência, as fases da mitose: Prófase, Prometáfase, Metáfase, Anáfase, Telófase e Citocinese. A respeito delas diga quais os processos intracelulares ocorrem em cada processo
Prófase – cromatina se condensa com o auxílio das condensinas, que contribuem para espiralar o DNA ao redor de sua estrutura; envelope nuclear ainda intacto; já são vistos dois pares de centríolos, cada um com um centrossomo, localizados em polos opostos da célula; começa a polimerização dos microtúbulos para a firmação do fuso mitótico.
Prometáfase – o envelope nuclear começa a se desfazer com a fosforilação das fibras de laminina da lâmina nuclear; fragmentação de organelas (como o retículo endoplasmático, mitocôndria e Complexo de Golgi, para que sejam segregados às futuras células-filhas); microtúbulos se conectam ao cinetócoro, no centrômero das cromátides.
Metáfase – formação do fuso mitótico, com o alinhamento de todos os cromossomos na “placa metafásica”, os quais também estão ligados aos microtúbulos através do cinetócoro; no último ponto de checagem é verificado se todos os cromossomos estão corretamente alinhados e conectados para que a célula possa passar para a próxima fase.
Anáfase – segregação dos cromossomos aos polos da célula, devido à clivagem das coesinas (o que “solta” as cromátides-irmãs, antes unidas) e à tração dos microtúbulos em direção a seu respectivo centrossomo.
Telófase – os cromossomos chegam aos polos, ficando próximos ao centrossomo; eles se descondensam; o envelope nuclear é remontado (as lamininas ligam-se novamente à medida que são desfosforiladas); formação do anel contráctil, formado por microfilamentos de actina, no meio da célula.
Citocinese – contração dos filamentos do anel contráctil, com a formação do sulco de clivagem, com a consequente divisão da célula ao meio, separando as duas células-filha.
10 – Embora o processo de meiose seja similar à mitose em certos pontos, os dois processos principalmente diferem quanto às etapas de divisão. Explique o processo de meiose, evidenciando as etapas semelhantes à mitose e as etapas de divisão celular.
O controle e a manutenção do ciclo celular são realizados através dos pontos de verificação e é a ligação das CICLINAS específicas com as CDK que indicam a passagem para próxima etapa, ou seja, a continuidade correta do ciclo. A Checagem de iniciação (G1/S) analisa se há condições ambientais para a célula se proliferar (nutrientes, fatores de crescimento), a Checagem da entrada em Mitose (G2/M) verifica se não há danos no DNA e a Checagem para a anáfase e citocinese se há problemas na formação do fuso mitótico e consequente problemas na segregação do DNA. Esse mecanismo funciona como um relógio que liga cada evento em um tempo específico e faz com que os mesmos sejam sempre ordenados. Como são acontecimentos cíclicos, essa verificação garante que cada evento ocorra somente uma vez e que as etapas sejam realmente finalizadas antes do inicio da próxima. Apesar de ser claramente um processo complexo e muitas vezes eficiente, alguns erros podem acontecer. É um processo facilmente adaptável à tipos celulares específicos ou condições ambientais adversas.
11 - Como são formadas as fibras do fuso mitótico/meiótico, o que as compõem, e quais os tipos de fibras? Como elas se ligam às cromátides?
As fibras do fuso, seja na meiose ou na mitose, são formadas por subunidades de tubulinas, as que compõem os microtúbulos. Estes são polimerizados a partir do centrossomo, onde são encontrados os centríolos. Os centríolos existem em pares na célula; durante ao ciclo celular, na fase S, eles se duplicam, mas permanecem próximos, em um só centrossomo. Com a progressão do ciclo, na prófase, durante a fase M, os centrossomos são duplicados e cada par de centríolos segue para um polo da célula. Dos dois pares saem várias projeções dos microtúbulos. São três tipos de fibras diferentes: os microtúbulos astrais, que seguem próximo à membrana plasmática e ficam em contato somente com ela; os microtúbulos, que conectam uma fibra do fuso à outra (de outro centrossomo), por meio de proteínas motoras (como cinesinas e dineínas); e os microtúbulos do cinetócoro, que se ligam ao cinetócoro das cromátides. Até a fase de prometáfase, as cromátides estão unidas pelo cinetócoro, uma estrutura proteica no centrômero do cromossomo que possui um “anel” por onde o microtúbulo pode se conectar e fixar-se. A fixação é importante para que, na metáfase, o cromossomo seja “partido” e as cromátides, segregadas, cada uma para um polo da célula, conforme os microtúbulos do cinetócoro “encurtam” em direção a seu respectivo centrossomo e os microtúbulos polares “alongam”, afastando os polos.
12 - Explique de que forma a célula consegue separar o material genético durante a última etapa de divisão celular.
As cromátides (ou cromossomos homólogos, na primeira fase da meiose), antes de se separarem, estão unidas entre si pelas coesinas e, no centrômero, estão ligadas a um complexo proteico, o cinetócoro, o qual possui uma estrutura por onde as fibras de microtúbulos do cinetócoro são capazes de se fixar. Ao entrar na metáfase, seja na mitose, meiose I ou meiose II, as coesinas são clivadas pelas separases (as quais são ativadas pela APC/C), possibilitando que o material genético seja separado. Os microtúbulos, que já estão conectados nos cinetócoros, despolimerizam e, com isso, encurtam, trazendo consigo a cromátide que foi dividida. Os cromossomos (meiose I) ou cromátides (mitose e meiose II), conectados ao microtúbulo, seguem em direção aos centrossomos, cada um em uma extremidade da célula. Quando a célula entra na telófase, o material genético chega ao polo da célula e o envelope nuclear volta a ser remontado. Começa a formação do anél contráctil, formado por microfilamentos de actina e associado a proteínas motoras (miosina). O anel fica no meio da célula em divisão, contraindo-se e formando uma fenda, até que a célula divide-se em duas células-filhas.
Morte Celular: Apoptose e Necrose
1 - Explique o que é o equilíbrio celular:
É a regulação do número de células em um determinado organismo ou tecido através do controle do processo de seu ciclo e morte celular, por meio de mecanismos moleculares presentes nas células. A regulação consiste no controle da proliferação (aumento na quantidade de células), diferenciação (capacidade das células em adquirir função específica) e morte celular (eliminação de células velhas, defeituosas ou não mais necessárias). Dessa forma, o número de células em um tecido ou órgão permanece sempre em um processo de equilíbrio dinâmico.
2 - Diga quais efeitos que a desregulação do equilíbrio celular pode causar na célula:
Defeitos nos mecanismos moleculares que mantêm o equilíbrio do tamanho de órgãos e tecidos nos organismos multicelulares podem levar à proliferação excessiva das células, aliado a pouca ou não ocorrência de diferenciação ou morte celular. Isso pode causar um acúmulo de células no tecido ou órgão. Também pode ocorrer morte excessiva, sem que haja proliferação das células, lesionando a região. A morte celular atua no desenvolvimento de órgãos, mantendo-o em seu tamanho correto, enquanto novas células surgem, e em sua forma ideal, como a degeneração da membrana interdigital do embrião, garantindo o formato dos dedos de mãos e pés; evita a permanência ou proliferação de células mutadas e desreguladas, que podem ocasionar em câncer; além disso, ela evita uma resposta imune exacerbada devido ao excesso de células de defesa ou por sua permanência do organismo. O excesso de morte celular pode levar a doenças neurodegenerativas ou piorar o processo de infarto. Nos dois casos podem ocorrer má formações no tecido lesionado.
3 - Cite quais são os dois principais tipos de morte celular e explique a principal diferença entre elas e cite exemplos de seus causadores:
Há dois principais tipos de morte celular: apoptose (principal tipo de morte celular programada) e necrose (acidental). A necrose não depende de energia para ocorrer; ela é causada por um rompimento da membrana permitindo a entrada de água e livre troca de íons com o meio extracelular. Isso faz com que a célula não consiga produzir energia de forma eficiente nem bombear íon de um lado a outro da membrana; consequentemente a célula rompe-se e entra em equilíbrio com o meio, podendo levar a lesões no tecido ou órgão. O processo de necrose leva a um processo de inflamação local. A apoptose, por outro lado, depende de energia para ocorrer; há ativação de um programa bioquímico muito bem definido de sinalização celular, levando a alterações morfológicas e bioquímicas dentro da célula. A célula se dissocia das células vizinhas e diminui de tamanho; o seu DNA se condensa e fragmenta-se; as organelas se dissociam e ocorre a formação de vesículas formadas por membrana, ou “blebs”, que formam os corpos apoptóticos. Moléculas presentes na face intracelular da membrana, como os fosfolipídeos de fosfatidilserina, sinalizam a células do sistema imune, como macrófagos e neutrófilos, que a célula apoptótica deve ser fagocitada e eliminada do organismo. Como o material intracelular não é disperso, não ocorre resposta inflamatória. Pode tratar-se de um processo fisiológico ou patológico no organismo logo, não necessariamente há lesões teciduais. Alguns fatores que causam a necrose são a falta de oxigênio, variações extremas de temperatura, agressões químicas ou físicas (ácidos, bases, choques, cortes e etc…) certos tipos de fármacos e agentes biológicos. Já a apoptose pode ocorrer devido a um processo fisiológico, como durante o desenvolvimento do feto, para formar tecidos e órgãos, mantendo-os em sua forma correta; ocorre para manutenção do tecido, com a morte de células velhas ou danificadas, junto à proliferação e diferenciação de células novas; atua em uma resposta imune eficiente e adequada, de forma que não seja exacerbada e danifique o organismo. Porém, a apoptose também pode ser causada como um processo patológico como infecção de vírus e bactérias; sua desregulação pode levar a doenças degenerativas ou tumores, dentre outras causas.
4 - Cite três outras formas de morte celular programada que não a apoptose, e explique-as, citando exemplos:
- Autofagia – é um processo de digestão de organelas e complexos moleculares no interior da células. Quando estrutura grandes na células se tornam velhas, obsoletas ou danificadas elas são envolvidas por uma dupla camada de membrana que se funde ao lisossomo, formando o autofagossomo. Todo o material digerido por esse processo pode ser reaproveitado pelas células; alguns tipos de tumores, drogas ou a escassez de nutriente levam à morte celular por disfunção de suas organelas;
- Netose – tipo de resposta imune, realizada por neutrófilos, células de defesa, que liberam seu material genético junto a agentes químicos e proteicos em forma de rede, fazendo com que patógenos (vírus, fungos, bactérias) fiquem aderidas nessa mistura e não contaminem o organismo;
- Cornificação – produção de queratina em excesso até que a membrana da célula seja rompida, perdendo sua integridade e liberando a proteína; processo responsável pela formação e impermeabilização dos epitélios queratinizados, cornos (em animais) e unhas.
5 - Qual a função do citocromo C no processo de Apoptose?
A liberação do citocromo C da mitocôndria no processo de apoptose pela via intrínseca induz a formação do Apoptossoma, que ativa a caspase 9 iniciadora e toda a cascata proteolítica da apoptose. A partir da saída do citocromo C da mitocôndria ele se encontra livre no citosol para poder se ligar e ativar o Apaf (Fator de Ativação das Proteases Apoptóticas) que expõem seu domínio CARD (Dominio de Recrutamento de Caspase). Esse domínio recruta e se une aos domínios de outros 6 Apafs ativos, essa nova formação com 7 Apafs unidas recruta e ativa 7 pro-caspases 9 através de seus domínios CARD formando o Apoptossoma. A aproximação das pró-caspases 9 no apoptossoma leva a sua autoproteolise e ativação. Caspase 9 ativa cliva e ativa a pró-caspase 3 efetora que levará ao fenótipo apoptótico.
6 - Como o citocromo C atravessa a membrana da mitocôndria no momento correto?
A mitocôndria possui em sua membrana externa diversas proteínas, porém, há um grupo especial denominado proteínas BH123 (formadoras de poro; pró-apoptóticas). Estas proteínas são ativadas pelas proteínas BH3-only que recebem estímulos apoptóticos que ativam ou aumentam sua expressão. Ao serem ativadas as proteínas BH123 se unem às proteínas BH123 levando a formação de um canal (poro na membrana externa da mitocôndria) que permite a passagem do citocromo de dentro da mitocôndria para o citosol. Entretanto, existem proteínas anti-apoptótica denominadas BH1234 que, estando ativas, são capazes de inibir a formação espontânea do poro pelas proteínas BH123. As proteínas BH3-only podem ser capazes de ativar a formação do poro e/ou de inibir a ação das proteínas anti-apoptótica BH1234, levando a liberação do citocromo C. Um balanço entre a ativação e expressão das proteínas BH3-only e BH1234 que regula se a células se manterá viva ou entrará em apoptose.
7 - DESCREVA as semelhanças e diferenças da via extrínseca e da via intrínseca da apoptose.
Inicialmente ambas as vias utilizam a mesma família de proteínas as Caspase (Cysteine protease aspatate specific) e evidentemente o objetivo das mesmas é a morte celular, porém, os sinais e alguns participantes do processo de cada via são distintos. A via extrínseca recebe sinas apoptóticos do meio exterior, através de sinais solúveis ou ligados a membrana celular de outras células. O sinal é transmitido pela membrana através dos chamados RECEPTORES DE MORTE que ativam as caspases iniciadoras 8 ou 10. Para ativar as respectivas caspases é necessário que a proteína adaptadora FADD faça uma “ponte” entre o receptor de morte e a Procaspase-8 ou 10. Esse Complexo Sinalizador Indutor de Morte, denominado DISC, aproxima as caspases iniciadoras que se auto-clivam (autoproteólise) levando a formação de hetero tetrâmero e à ativação dessas caspases, estimulando toda a cascata proteolítica. Sinas completamente distintos da via extrínseca ativam a via intrínseca, como o nome induz são sinais internos, como por exemplo danos no DNA, ausência de fator de crescimento etc. Estes sinas provocam a liberação de citocromo C pela mitocôndria, que leva a formação do apoptossoma e a ativação da caspase 9 iniciadoras e toda a cascata proteolítica. As caspases iniciadoras (8, 9 e 10) proteolisam e ativam as caspases efetoras (principlamente caspase 3) que irá induzir o fenótipo apoptótico na célula.