Morte Celular: Apoptose e Necrose
Slides da Aula
FAQs em Vídeos
Estudo Dirigido
Testando Seu Conhecimento
Vídeos
Clicando no botão abaixo você encontrará diversos vídeos sobre o tema.
![]()
Imagens
Esta página também contém imagens que vão auxiliar no seu aprendizado. Aproveite!
Figuras Legendadas

Fig. 1 O equilíbrio celular. A manutenção do número de células em um organismo é altamente regulada por mecanismos presentes em todos os seres multicelulares. As células de tecidos e órgãos, por exemplo, param de proliferar quando essas estruturas atingem seu tamanho específico, e suas células antigas se renovam em certos períodos de tempo (dependendo do tecido e suas células, podem se renovar constantemente, raramente ou nunca se renovarem); células do sistema imune podem proliferar o suficiente para eliminar um micro-organismo, mas não para causar danos ao corpo pois após eliminação do micro-organismo as células do sistema imune morrem (com exceção de certas doenças autoimunes). Para que o organismo permaneça vivo, é necessário que haja proliferação, diferenciação e morte celular adequadas e balanceadas, como mostrado na figura acima. A proliferação é a entrada no ciclo celular (o aumento do número de células) e é fundamental para o desenvolvimento, crescimento, regeneração e para manter o organismo vivo. A diferenciação celular é a capacidade das células em adquirir função específica, de acordo com seu órgão, conforme as células proliferam; a diferenciação leva a parada do ciclo celular, levando a um equilíbrio com a proliferação celular. Por fim, a morte celular mantém o número de células sempre constante, eliminando células velhas ao mesmo tempo que surgem células novas ou que estas se diferenciem.

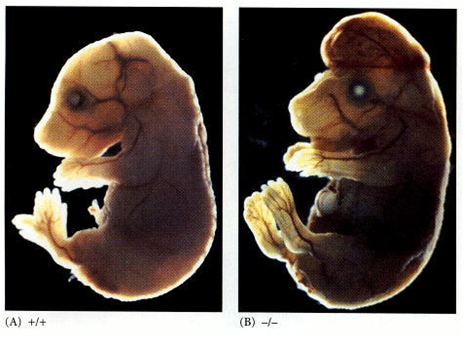
Fig. 2 Desregulação do Equilíbrio Celular. Quaisquer defeitos nos mecanismos moleculares que regulam as etapas do ciclo e morte celular podem desregular o equilíbrio celular. Uma das consequências disso é o excesso de proliferação quando há pouca ou não há diferenciação ou morte celular (ilustrado na figura A), levando ao acúmulo de células no tecido ou órgão. A morte celular é fundamental para o desenvolvimento de órgãos, mantendo-o em seu tamanho correto, enquanto novas células surgem, e em sua forma ideal, como a degeneração da membrana interdigital do embrião, garantindo o formato dos dedos de mãos e pés (D). É importante para evitar a permanência ou proliferação de células mutadas, que podem ocasionar em câncer (figura C, mostrando um melanoma, também conhecido como câncer de pele) ou para evitar uma resposta imune exacerbada devido ao excesso de células de defesa ou por sua permanência do organismo. Outra consequência, pelo contrário, é a falta de proliferação enquanto há diferenciação e morte celular, sejam estas ocorrendo em seu período normal ou em excesso (ilustrado na figura B). Isso pode levar a doenças neurodegenerativas (a figura E mostra, à esquerda, como fica evidente a degeneração de um cérebro, em comparação com um cérebro saudável, à direita) ou piorar o processo de infarto. O acúmulo de células pode dar origem aos diversos tipos de câncer, doenças autoimunes e más formações.

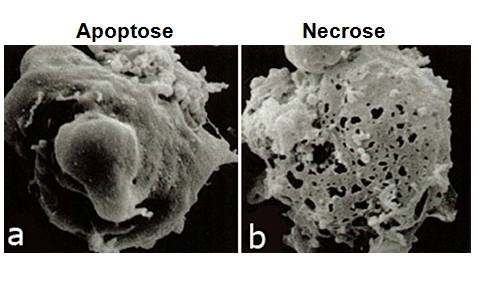
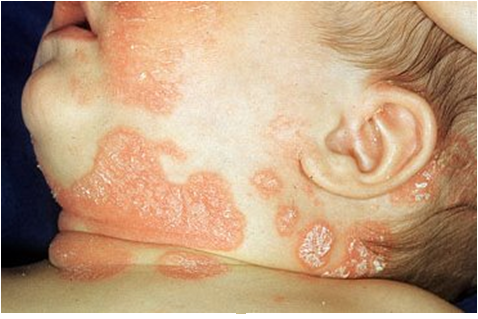
Fig. 3 Tipos de Morte Celular: Necrose e Apoptose. A morte celular pode ocorrer de forma acidental (ou necrose), ou de forma programada (ou, geralmente, apoptose). A Necrose não depende de energia para ocorrer; a membrana da célula perde sua integridade e rompe-se, fazendo com que entre água em seu interior e terminando por causar seu inchaço, ruptura e equilíbrio com o meio. Entre os causadores da necrose estão a falta de Oxigênio (dificultando ou impedindo a produção de energia na célula); agentes físicos, como altas ou baixas temperaturas e radiação; agentes químicos (como fármacos) e biológicos (como vírus, bactérias ou fungos; caso da figura E). O outro tipo de morte celular, a Apoptose, é o processo mais comum de morte celular programada e ocorre com gasto de energia por parte da célula que sofrerá a morte celular. Há ativação de um programa bioquímico definido de sinalização celular, levando a alterações morfológicas e bioquímicas na célula. É um processo que ocorre naturalmente no organismo, que serve para manter tecidos e órgãos em seus devidos tamanhos, definindo seu limites (como na metamorfose do girino e retração de sua cauda por apoptose, figura A; e na formação dos dedos através da degeneração e retração da membrana interdigital, figura B); a figura C mostra quando ocorre falha no processo de apoptose o que pode causar a permanência completa ou parcial da membrana interdigital na criança). Serve também à resposta imune (tornando-a eficiente, com início e término adequados, e não de forma exacerbada ou contínua, para não prejudicar o organismo), na diferenciação terminal (quando a célula se diferencia e, ao final, morre por apoptose) e, inclusive, na eliminação de células anormais (seja por estresse ou dano no DNA). O processo de apoptose está associado à eliminação de células infectadas; de células do sistema imune quando uma resposta não é mais necessária (quando o micro-organismo já foi eliminado do corpo); de células mutadas, com danos no DNA, células que possuem receptores não funcionais ou que perderam a função; de células em excesso (tanto para conformação do tecido como para encerrar a resposta imune); desnecessárias (relativo ao desenvolvimento do organismo) ou células velhas. É importante ressaltar que a apoptose é apenas um tipo de morte celular programada; entre outros estão a necroptose (necrose programada), a autofagia (processo de degradação de organelas e grandes estruturas celulares, podendo levar a sua morte; principalmente ligado a carência de nutrientes), a netose (realizada por neutrófilos, células de defesa) e a cornificação (produção de queratina em excesso até que a membrana da célula se rompa, feito por células epiteliais, por exemplo). A figura D mostra a diferença na superfície de uma célula normal e em processo de apoptose, apresentando as diversas vesículas de membrana (chamadas de blebs de membrana). Posteriormente, essas vesículas se soltam da célula para formar corpos apoptóticos que contém pedaços de organelas e DNA e que impedem que esse material e o material genético se dispersem pelo tecido; a figura E mostra a diferença entre a célula em apoptose e em necrose, ressaltando os diversos “furos” presente na membrana, conforme ela se rompe.

Fig. 4 Características morfológicas da necrose e da apoptose. Na necrose, devido ao estresse osmótico a célula perde a integridade de sua membrana e rompe-se aos poucos. Por causa de seu gradiente osmótico e eletroquímico (a célula é mais concentrada que seu meio), a água extracelular tende a entrar na célula e em suas organelas. A célula tende a ficar em equilíbrio com o meio e, além disso, ela ganha volume até se romper. A célula não consegue exercer suas funções, como produção de ATP e bombeamento de íons e água para o exterior da célula, e acaba se deteriorando (D). Um dos causadores da necrose é a falta de Oxigênio para as células: sem Oxigênio, elas não conseguem carrear os elétrons necessários para a produção de energia da célula; a produção energética diminui e as funções da célula cessam. Agentes físicos também levam à necrose, como temperaturas muito altas (fazendo com que a célula perca água por evaporação), ou muito baixas, congelando-as e formando cristais dentro da célula que rasgam a membrana (após “descongelarem”, os cristais somem, mas a membrana permanece rompida). Outros agentes, como químicos (fármacos) e biológicos (vírus, bactérias ou fungos) podem causar o rompimento da membrana da célula alvo, sem ativar programas bioquímicos pré-definidos (característico da apoptose). O processo de necrose acaba deflagrando uma resposta inflamatória devido ao material intracelular que se espalha no meio, como proteínas, enzimas, nucleotídeos (como o material genético não se condensa, ele se dispersa); isso atrai células do sistema imune, como neutrófilos, macrófagos e linfócitos, para o local onde está ocorrendo uma suposta invasão, seja causada por vírus, fungos ou infecção bacteriana. A figura A mostra a diferença entre uma célula normal e outra em processo de apoptose.
Por outro lado, na apoptose, enzimas e mecanismos moleculares atuam em um programa bioquímico definido de sinalização celular, levando a alterações morfológicas e bioquímicas na célula; a célula se dissocia das células vizinhas e diminui de tamanho, ocorre condensação e fragmentação do material genético (figura B, apontado pelas setas; o DNA está em azul), dissociação das organelas e formação de vesículas formadas por membrana, chamadas blebs, que formam corpos apoptóticos (podem ser visto na figura A, como as vesículas na superfície mais azuladas, e esquematizado em E). Dessa forma, não há dispersão do material genético e, portanto, nenhuma resposta inflamatória é desencadeada; trata-se de um processo que pode ser comum a todas as células como, por exemplo, para a manutenção do tamanho do órgão ou tecido, ou devido a um processo infeccioso, dano celular, problemas na diferenciação celular e outros. Além disso, moléculas presentes na face intracelular da membrana (os fosfolipídeos de fosfatidilserina, destacadas em verde na figura C) são expostas (passam para a face extracelular), sinalizando para células do sistema imune, como macrófagos, que a célula deve sofrer fagocitose e ser rapidamente retirada do tecido.

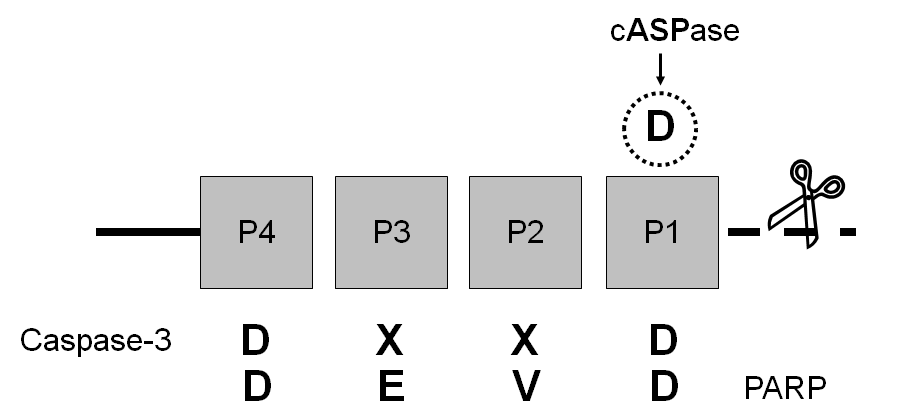
Fig. 5 Família de proteases das caspases. A Caspase (Cysteine protease aspatate specific) é a principal proteína responsável pelo processo de apoptose. Cada membro da família possui um sítio de clivagem específico que determina sua proteína alvo. Como mostrado em A, as caspases são sintetizadas na forma de pró-caspases inativas, e são ativadas pelo proteólise (por isso são zimogênios). Quando duas pró-caspases são aproximadas físicamente elas se clivam liberando o domínio pró e separando a unidade catalítica em duas subunidades, subunidade grande e pequena (A). Duas subunidades pequenas e duas grandes se oligomerizam (se juntam) para formar um heterotetrâmero de caspase agora completamente ativo. As Caspases são dividias funcionalmente em Caspases Iniciadoras (8, 9, 10) e Caspases Efetoras (3 principalmente). As Caspases iniciadoras representadas em B, estimulam e iniciam a cascata proteolítica da apoptose. As caspases iniciadoras reconhecem um sinal de morte celular através de seu Domínio CARD (caspase recruitment domain; Caspase 9) ou do Domínio DED (death effector domain; Caspases 8 e 10), ambos domínios de interação proteína-proteína, que possibilitam a aproximação dessas proteínas no inicio do processo de apoptose. As Caspases efetoras representadas em C, são responsáveis pela degradação das proteínas celulares alvo que levarão ao fenótipo apoptótico (D). São inúmeros os alvos das caspases efetoras, como a lâmina nuclear que provoca a dissolução do núcleo, proteínas de adesão celular e remodelamento do citoesqueleto que induz a diminuição da célula e sua fragmentação, exposição de sinais de “coma-me” (levando a exposição do fosfolipídeo de fosfatidilserina), inibição da tradução etc.

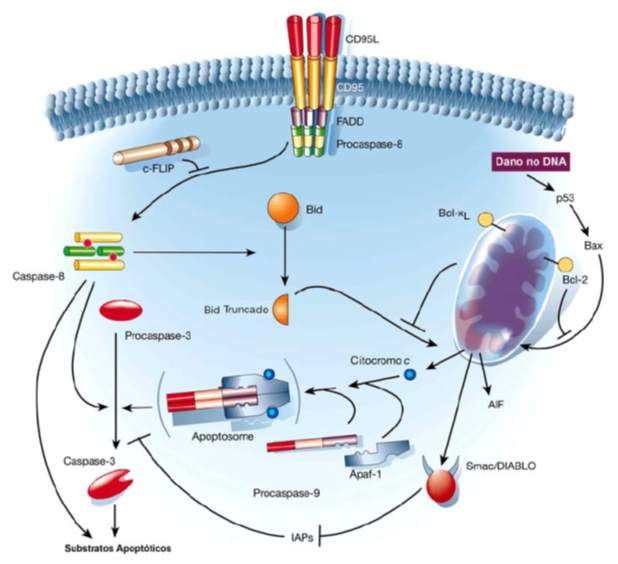
Fig. 6 Via extrínseca de apoptose. A imagem representa todo o mecanismo de Apoptose através de sinais solúveis ou ligados a membrana celular, como é o caso do Linfócito representado, provenientes do meio extracelular (Fas,TNF, TRAIL). O sinal é transmitido pela membrana através dos chamados RECEPTORES DE MORTE que ativam as caspases iniciadoras 8 ou 10. Para ativar as respectivas caspases é necessário que a proteína adaptadora FADD faça uma “ponte” entre o receptor de morte e a Procaspase-8 ou 10. Esse Complexo Sinalizador Indutor de Morte, denominado DISC, cliva e ativa as caspases iniciadoras através de sua aproximação e auto-proteólise, estimulando toda a cascata proteolítica.

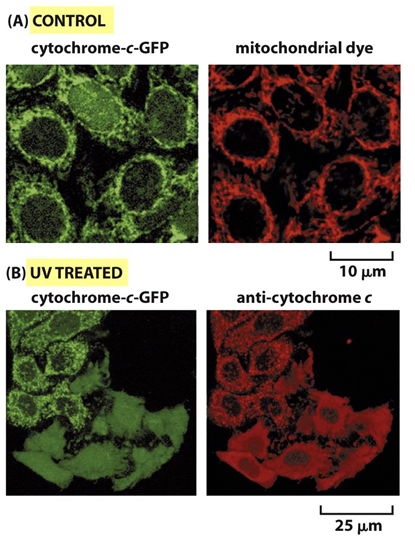
Fig. 7 Via intrínseca de apoptose. A imagem representa a via extrínseca da apoptose vista anteriormente (A) e a via intrínseca de apoptose (B). Como esquematizado, a via intrínseca se inicia na mitocôndria com a liberação de citocromo C que leva a formação do apoptossoma e a ativação da caspase iniciadora 9. Sinais completamente distintos da via extrínseca ativam a via intrínseca, como o nome induz são sinais internos, como por exemplo danos no DNA, ausência de fator de crescimento etc. Mas a membrana da mitocôndria é extremamente impermeável para permitir seu próprio funcionamento, como é possível a saída de citocromo C de seu interior? É isso que veremos na próxima figura.

Fig. 8 Via intrínseca de apoptose: permeabilização da mitocôndria. Como representado em A a mitocôndria possui uma membrana extremamente impermeável para a maioria das substâncias ao seu redor, entretanto , ela apresenta diversas proteínas transmembrana em sua estrutura. O estímulo apoptótico intrínseco induz a ativação de algumas destas proteínas, denominadas proteínas BH123 (proteínas pró-apoptóticas formadoras de canal), que formam um canal em sua forma ativa liberando o citocromo C de dentro da mitocôndria para o citosol, como mostrado em B. Porém, se uma proteína antiapoptótica da família BH1234 (exemplificado pela proteína Bcl2 na figura) estiver ativa ela possui capacidade de inibir a formação do canal pelas proteínas BH123,representada na figura B e C. Os estímulos apoptóticos ativam ou aumentam a expressão das proteínas BH3-only (proteínas pró-apoptóticas). Essas proteínas (BH3-only) são capazes de ativar diretamente a formação do canal pelas proteínas BH123 ou de inibirem a atividade das proteínas anti-apoptóticas (BH1234), levando portanto a formação do canal e liberação do citocromo C. Sinais que levam a ativação ou expressão das proteínas BH3-only são: dano no DNA (ativação p53) – Puma, Noxa, Bid; ausência de substrato sólido de adesão – Bim1, BMF; ausência de fator de crescimento – Bad, Bim1, dentre outros.

Fig. 9 Via intrínseca de apoptose: Apoptossoma. A imagem representa todos os processos que precisam ocorrer para ativar a caspase 9 iniciadora na via intrínseca a partir da saída do citocromo C da mitocôndria. O Apaf (Fator de Ativação das Proteases Apoptóticas) é ativado pelo citocromo C, expondo seu domínio CARD (Dominio de Recrutamento de Caspase) que se une aos domínios de outros 6 Apafs ativos, como representado na figura. Essa nova formação com 7 Apafs unidas recruta e ativa 7 pro-caspases 9 (através do seu domínio de interação CARD). A ativação da pró-caspase 9 se dá por sua aproximação pela formação do apoptossomo. Uma vez aitva, a caspase 9 ativa a cascata proteolítica ativando as Caspase 3 efetora e todo o mecanismo de apoptose.
Imagens Complementares
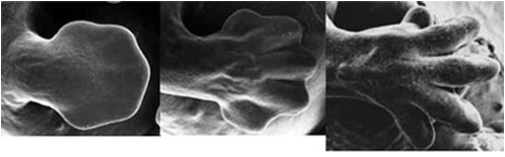
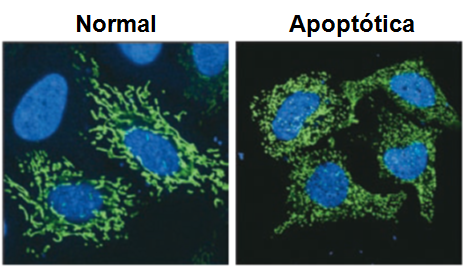
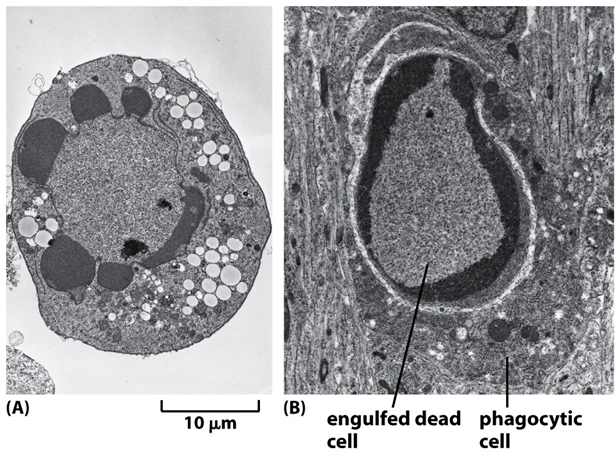
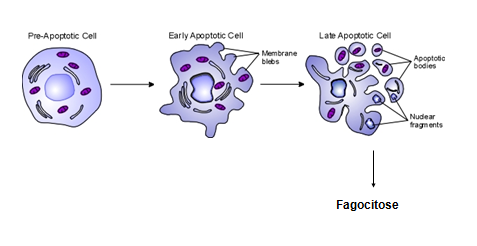
Todas as imagens foram retiradas da seguinte fonte: (c) 2014 from Molecular Biology of the Cell, Sixth Edition by Alberts et al. Reproduced by permission of Garland Science/Taylor & Francis Group LLC.