Mitocôndrias e Peroxissomos
Slides da Aula
Mapa Conceitual
FAQs em Vídeos
Estudo Dirigido
Testando Seu Conhecimento
Aula Gravada
Vídeos
Clicando no botão abaixo você encontrará diversos vídeos sobre o tema.
![]()
Imagens
Esta página também contém imagens que vão auxiliar no seu aprendizado. Aproveite!
Figuras Legendadas

Fig. 1 Peroxissomos. Todas as células possuem Peroxissomos, que são organelas simples com apenas uma membrana lipídica. Suas principais funções são detoxificação da célula (etanol, fenóis, ácido fórmico, formaldeído) e b-oxidação (produção de Acetil-CoA pela oxidação de ácidos graxos). Em seu interior possuem oxidases que removem átomos de hidrogênios de substratos orgânicos em reações oxidativas, produzindo peróxido de hidrogênio (H2O2). A H2O2 é utilizada para oxidar e eliminar essas substâncias toxicas da célula, em uma reação metabolizada pela enzima catalse. Apesar de ser uma reação extremamente importante para o funcionamento celular a água oxigenada (H2O2 ) pode causar lesões em sua estrutura. O excedente de água oxigenada pode ser quebrada em água e gás oxigênio também pela enzima catalase.

Fig. 2 Proteínas do Peroxissomo. Como observamos na imagem, todas as proteínas do Peroxissomo são importadas diretamente do citoplasma. Proteínas que devem ser importadas para o Peroxissomo possuem uma Sequencia sinal de importação para o peroxissomo. Resumidamente, essas proteínas são traduzidas pelo Ribossomo no citoplasma. A sequencia Sinal é reconhecida pelo receptor de sequencia sinal de importação para o peroxissomo a encaminhando para o translocador de proteínas, presente na membrana desta organela. O translocador catalisam a importação da proteína através da membrana do Peroxissomo. São estas proteínas, de origem citoplasmática, que executam todas as funções essências desta organela.

Fig. 3 Origem do Peroxissomo. Como observamos na imagem, os Peroxissomos se originam no Reticulo Endoplasmático, através de vesículas precursoras. Estas vesículas apresentam proteínas específicas que catalisam a importação de proteínas e lipídeos do citosol (translocadores). A vesícula cresce devido a captação destes constituintes do citosol, que também provem sua diferenciação e aquisição de funções específicas.


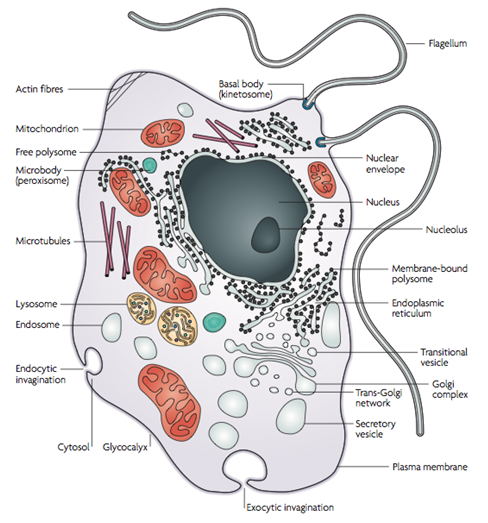
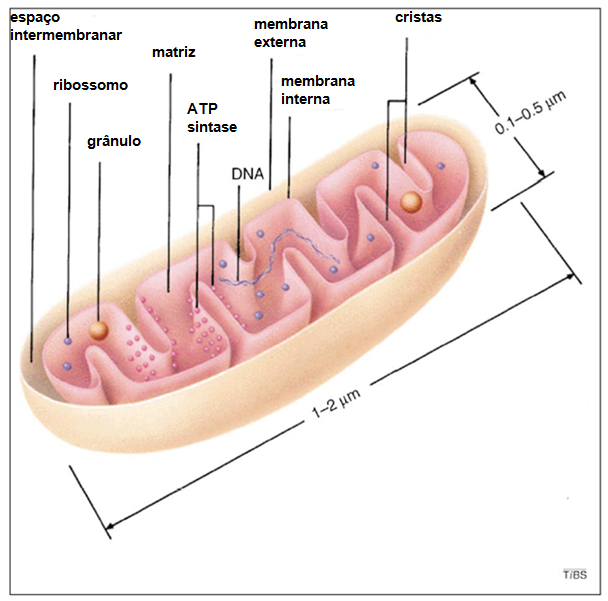
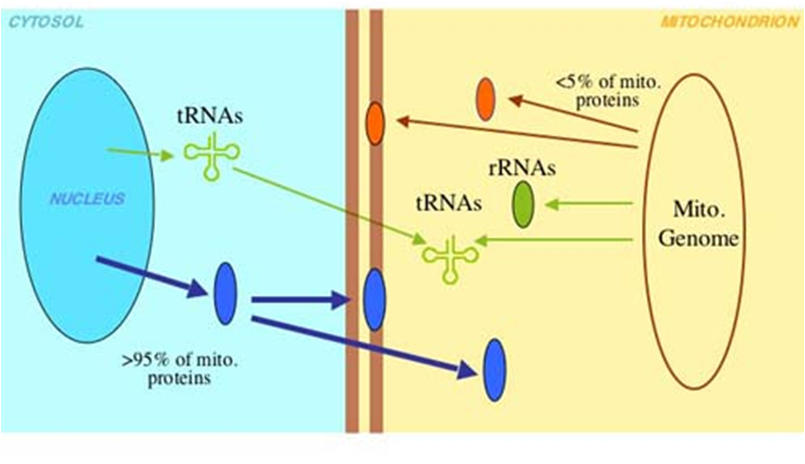
Fig. 4 Mitocôndria. Em (A) observamos uma microscopia eletrônica da Mitocôndria, uma organela complexa e de extrema importância para a célula. A mitocôndria possui função principal de produção de ATP através da oxidação de acetil-CoA, originado da glicose (glicolise), de lipídeos (b-oxidação) ou proteínas. Em (B) estão destacadas todas suas divisões e estrutura. A mitocôndria possui dois espaços, o mais interno que chamamos de matriz mitocondrial e o espaço intermembrana. Estes espaços são essências, pois são eles que garantem o grande gradiente eletroquímico de hidrogênio que possibilita a produção de ATP pela ATP sintase. Na matriz ocorre o Ciclo de Krebs e estão localizados todos os ácidos nucléicos da organela, incluindo seu DNA circular. A membrana externa possui porinas e complexos transportadores (TOM). A membrana interna é mais fluída (ausência de colesterol) e menos permeável (presença de cardiolipina), aspectos importantes para a manutenção do gradiente eletroquímico de H+. Devido a abundância de ATP sintases, cadeias transportadoras de elétrons e alguns complexos transportadores (TIM), a membrana interna tem a maior relação proteína/lipídeo da célula (70/30).

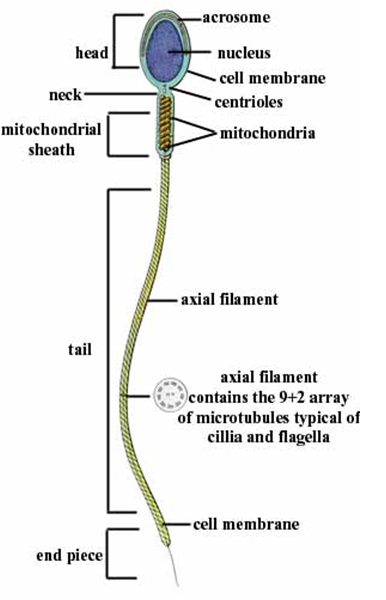
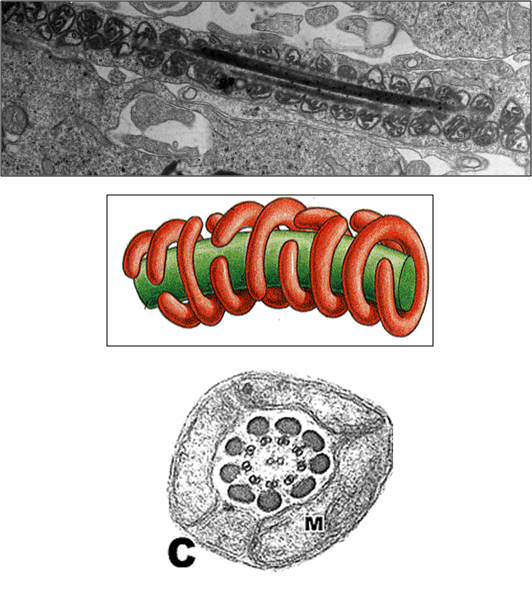
Fig. 5 Localização subcelular estratégica da Mitocôndria. A Mitocôndria, em células especializadas, tem a tendência de se localizar próxima a regiões de grande demanda energética. Como por exemplo, na imagem, em que Mitocôndrias estão intimamente relacionadas às miofibrilas, que possuem uma grande necessidade de ATP para executar o processo de contração. Assim como nos espermatozoides, onde estão próximas à cauda, que necessita de muito ATP para realizar seu movimento.

Fig 6. Origem dos substratos para a produção de ATP. Normalmente o substrato utilizado é a glicose, como fonte rápida de energia, obtida da alimentação ou da quebra do glicogênio e, em certos casos, os ácidos graxos dos lipídeos encontrados em adipócitos. Como mostrado na figura A, uma molécula de glicose (de seis carbonos) pode ser transformada através da glicólise em duas moléculas de piruvato (de três carbonos cada uma), presentes no citosol; essa reação gera um saldo líquido de 2 ATPs e 2 NADH + H+ e ocorre independentemente se as vias seguintes serão a aeróbica ou a anaeróbica. Na respiração aeróbica, o piruvato é encaminhado à mitocôndria, sendo oxidada pelo complexo piruvato desidrogenase e transformada em Aceti-Coa (com 2 Carbonos). Já os ácidos graxos (figura B), na matriz mitocondrial ou no peroxissomo, sofrem β -oxidação, ou seja, uma clivagem a cada dois Carbonos em sua cauda de hidrocarbonetos, que pode variar em tamanho, que termina por gerar no mínimo 5 moléculas de Acetil-Coa, NADH + H+ (e FADH2, não mostrado na figura). No final, gera várias moléculas de ATP (uma única molécula de ácido palmítico, por exemplo, possui 16 carbonos e pode, portanto, gerar um saldo líquido de 106 ATPs no final da respiração aeróbica). Ambos Acetil-Coa dos dois substratos podem ser utilizados na fase seguinte da respiração celular, o Ciclo de Krebs.

Fig 7. Ciclo de Krebs e Cadeia Respiratória. São, respectivamente, a segunda e terceira fases da respiração celular aeróbica (com uso de Oxigênio), sendo a glicólise a primeira fase. O Acetil-CoA proveniente da glicose ou da β-oxidação de ácidos graxos entra no ciclo de Krebs (ou Ciclo do ácido cítrico), reagindo com o oxaloacetato pré-existente do ciclo e formando o ácido cítrico. Este, ao longo da via, sofrerá uma série de reações que irão desencadear na formação de um novo oxaloacetato, tendo como saldo líquido a formação de 3 NADH + H+, que permanecem no citoplasma, e um CO2, que será expelido na respiração. O NADH funciona como um aceptor temporário de elétrons; seu elétron altamente reativo em sua forma reduzida será utilizado na cadeia transportadora de elétrons, ou cadeia respiratória, para a geração de um gradiente eletroquímico de H+ na membrana interna da mitocôndria para posterior produção de ATP pela ATP sintase.

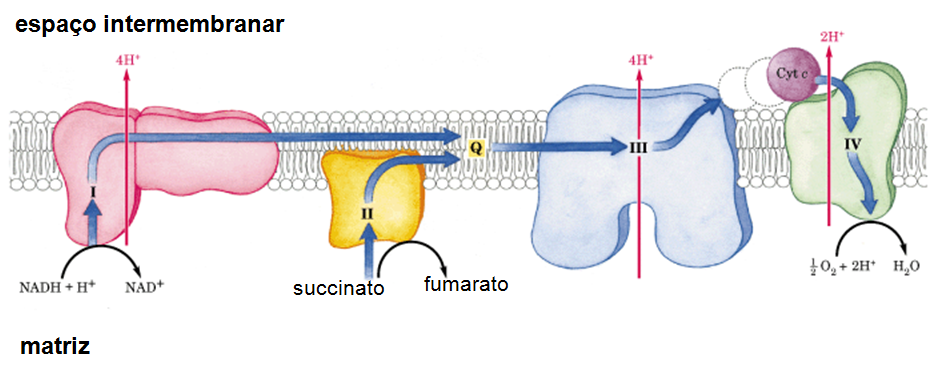
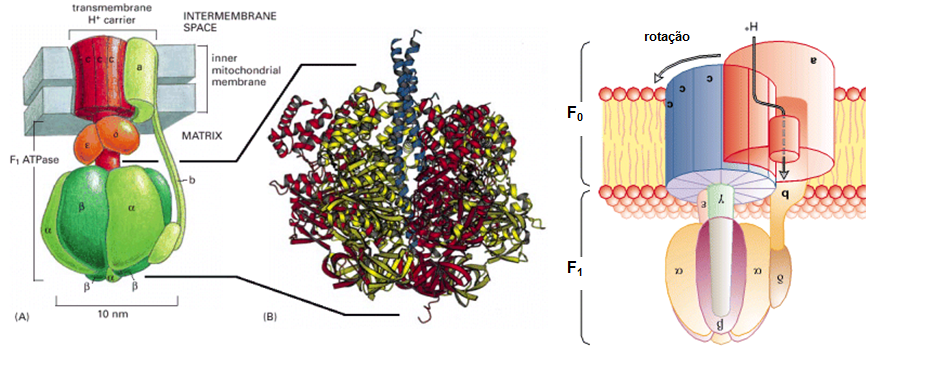
Fig 8. Cadeia transportadora de elétrons (ou cadeia respiratória). É composta por quatro complexos enzimáticos (sendo que o 1º, o 3º e o 4º são transmembranares e bombas de H+), presentes na membrana interna da mitocôndria. O complexo I (Complexo NADH Desidrogenase) é capaz de remover o H do NADH + H+ (proveniente do ciclo de Krebs) e, com isso, remover seu elétron energicamente carregado. Este elétron é transferido através de uma ubiquinona para o complexo III (Complexo Citocromo B-C1), e deste para o complexo IV (Complexo Citocromo Oxidase), através do citocromo C. Este último complexo é responsável por transferir o elétron para seu aceptor final, o O2 (proveniente da respiração), formando H2O, que permanece na célula. Quando o elétron transita pelos complexos, estes sofrem alterações conformacionais que promovem o bombeamento de prótons H+ da matriz mitocondrial para o espaço intermembranar; os complexos I e III bombeiam 4 H+, o IV bombeia 2H+ e o II, por não ser transmembrana, não bombeia elétrons (apenas o transfere ao complexo III, via ubiquinona). A transferência dos prótons torna o espaço intermembranar não apenas mais concentrado em H+, mas também o torna mais ácido (pH 7, em contraste ao pH 7,5 da matriz) e deixa a face da membrana voltada a ele mais positiva do que a face voltada à matriz, formando então um gradiente eletroquímico do H+, que tende a voltar à matriz. Como a membrana interna da mitocôndria é impermeável a íons, o H+ não a atravessa livremente, mas utiliza a ATPsintase (ou F0F1 ATPsintase), bomba do tipo F localizada na mesma membrana. Os prótons de H+ passam por seu canal e, com isso, causam uma mudança de conformação em sua subunidade, que é capaz de “girar” e promover, na matriz mitocondrial, a síntese de ATP a partir do ADP e do Pi presentes na matriz.

Fig. 9 Gradiente Eletroquímico na membrana interna da mitocôndria. O gradiente é gerado através do bombeamento de vários prótons de H+ da matriz para o espaço intermitocondrial, e possui dois componentes: o elétrico, composto pelo potencial de membrana, que torna a face voltada ao espaço intermembranar mais positiva do que a voltada à matriz, devido à carga positiva dos íons de Hidrogênio (pela polaridade, a tendência do H+ é voltar à matriz); e o químico, representado pelo gradiente de concentração dos íons H+ que torna-se maior no espaço intermembranar do que na matriz (logo, a tendência do H+ é também voltar à matriz, seguindo seu gradiente). Essas duas forças formam, então, um gradiente eletroquímico, e a tendência do próton é retornar à matriz.

Fig. 10 Sequência sinal da proteína direcionada à mitocôndria e translocação da proteína para a matriz mitocondrial feita por translocadores proteicos. Todas as proteínas são importadas diretamente do citoplasma; uma sequência sinal é traduzida junto à proteína, que é reconhecida por um complexo proteico que catalisa a importação, localizado na membrana externa, chamado de Complexo TOM. Este possui um receptor para a sequência sinal da proteína, capaz de reconhecê-la e direcioná-la ao canal de translocação mitocondrial. A proteína, então, atravessa a membrana externa e é capaz de ficar no espaço intermembranas ou pode ser reconhecida por outro complexo, o Complexo TIM, na membrana mitocondrial interna, que logo capta a proteína e a atravessa por seu canal, inserindo-a na matriz mitocondrial. Na matriz, a proteína terá sua sequência sinal clivada por peptidases-sinal e adquirirá sua conformação, tornando-se uma proteína madura. A figura A mostra o arranjo dos Complexos TOM e TIM nas membranas mitocondriais, e a figura B demonstra como ocorre essa translocação para a matriz.
Imagens Complementares